कंकाल की मांसपेशी लिपिड घुसपैठ, जिसे मायोस्टेटोसिस के रूप में जाना जाता है, मोटापे और उम्र बढ़ने के साथ बढ़ता है। मायोस्टेटोसिस मांसपेशियों और ताकत के साथ और इंसुलिन संवेदनशीलता के साथ नकारात्मक रूप से सहसंबद्ध है1. इसके अलावा, हाल के अध्ययनों से संकेत मिलता है कि मायोस्टेटोसिस की डिग्री का उपयोग अन्य स्थितियों जैसे हृदय रोग2, गैर-अल्कोहल फैटी यकृत रोग 3, या कैंसर4 के लिए पूर्वानुमान कारकके रूप में किया जा सकता है। लिपिड मांसपेशियों के तंतुओं के बीच कंकाल की मांसपेशियों में एक्स्ट्रामायोसेल्युलर लिपिड के रूप में या फाइबर के भीतर इंट्रामायोसेल्युलर लिपिड (आईएमसीएल) के रूप में जमा हो सकते हैं। आईएमसीएल मुख्य रूप से लिपिड बूंदों (एलडी) में ट्राइग्लिसराइड्स के रूप में संग्रहीत होते हैं जो शारीरिक व्यायाम 5,6 के दौरान चयापचय ईंधनके रूप में उपयोग किए जाते हैं। हालांकि, जब लिपिड की आपूर्ति मांग से अधिक हो जाती है, या जब माइटोकॉन्ड्रिया बेकार हो जाता है, तो आईएमसीएल को मांसपेशियों के इंसुलिन प्रतिरोध में फंसाया जाएगा, जैसा कि चयापचय रूप से अस्वास्थ्यकर, मोटापे से ग्रस्त व्यक्तियों और टाइप 2 मधुमेह रोगियों में देखाजाता है। दिलचस्प बात यह है कि धीरज एथलीटों में उच्च इंसुलिन संवेदनशीलता बनाए रखते हुए टाइप 2 मधुमेह मेलिटस वाले मोटापे से ग्रस्त रोगियों में पाए जाने वाले आईएमसीएल के स्तर समान हैं, यदि उच्च नहीं हैं। इस घटना को “एथलीट के विरोधाभास” 8,9 के रूप में वर्णित किया गया है, और मांसपेशियों के एलडी के अधिक सूक्ष्म मूल्यांकन द्वारा समझाया गया है, जो उनके आकार, घनत्व, स्थानीयकरण, गतिशीलता और लिपिड प्रजातियों की संरचना से संबंधित है।
सबसे पहले, एलडी आकार इंसुलिन संवेदनशीलता और शारीरिक फिटनेस10,11 से विपरीत रूप से सहसंबद्ध है। वास्तव में, छोटे एलडी लाइपेज कार्रवाई के लिए अपेक्षाकृत अधिक सतह क्षेत्र प्रदर्शित करते हैं और इस प्रकार, संभावित रूप से लिपिड12 को जुटाने की अधिक क्षमता रखते हैं। दूसरा, एलडी घनत्व (संख्या / सतह) इंसुलिन कार्रवाई 8,10 में एक विवादास्पद भूमिका निभाता है; फिर भी, यह एथलीटों में वृद्धि हुई प्रतीत होती है। तीसरा, एलडी का उपकोशिकीय स्थानीयकरण महत्वपूर्ण है, क्योंकि सतह झिल्ली (सबसरकोलेम्मल या परिधीय) के ठीक नीचे स्थित एलडी केंद्रीय लोगों 8,9,13 की तुलना में इंसुलिन संवेदनशीलता पर अधिक हानिकारक प्रभाव डालते हैं। उत्तरार्द्ध केंद्रीय माइटोकॉन्ड्रिया को ईंधन प्रदान करता है, जिसमें अधिक श्वसन गतिविधि होती है और संकुचन14 के लिए आवश्यक उच्च ऊर्जा मांग को पूरा करने के लिए अधिक विशिष्ट होते हैं। इसके विपरीत, परिधीय एलडी सबसरकोलेमल माइटोकॉन्ड्रिया की आपूर्ति करते हैं, जो झिल्ली से संबंधित प्रक्रियाओं में शामिलहोते हैं 8. अंत में, ट्राइग्लिसराइड्स से परे, मांसपेशियों के भीतर विशिष्ट जटिल लिपिड दूसरों की तुलना में अधिक हानिकारक हो सकते हैं। उदाहरण के लिए, डायसिलग्लिसरॉल, लंबी श्रृंखला एसाइल-सीओए और सेरामाइड्स मांसपेशियों में जमा हो सकते हैं जब ट्राइग्लिसराइड टर्नओवर दर कम होती है, जिससे इंसुलिन सिग्नलिंग 9,15 खराब हो जाती है। “एथलीट के विरोधाभास” पर लौटते हुए, धीरज एथलीटों में टाइप I (ऑक्सीडेटिव) फाइबर में ऊंची टर्नओवर दर के साथ छोटे केंद्रीय एलडी की एक उच्च संख्या होती है, जबकि मोटापे से ग्रस्त और मधुमेह रोगियों में टाइप II (ग्लाइकोलाइटिक) फाइबर 8,15,16 में कम टर्नओवर दरके साथ बड़े परिधीय एलडी होते हैं। ऊर्जा भंडारण और रिलीज में उनकी भूमिका के अलावा, व्युत्पन्न फैटी एसिड (एफए) और एक कोट प्रोटीन (पेरिलिपिन 5) के माध्यम से एलडी एफए ऑक्सीकरण और माइटोकॉन्ड्रियल बायोजेनेसिस8 के ट्रांसक्रिप्शनल विनियमन में शामिल महत्वपूर्ण खिलाड़ियों के रूप में भी कार्य कर सकते हैं। शरीर विज्ञान और पैथोफिजियोलॉजी में उनके महत्वपूर्ण निहितार्थों के कारण, एलडी गतिशीलता और कार्यों पर गहन अध्ययन की आवश्यकता है।
यद्यपि आईएमसीएल का अध्ययन करने के लिए कई तकनीकें हैं, वे फाइबर-विशिष्ट तरीके से एलडी आकार, घनत्व और वितरण को सटीक रूप से मापने के लिए उपयुक्त नहीं हैं। उदाहरण के लिए, चुंबकीय अनुनाद स्पेक्ट्रोस्कोपी द्वारा आईएमसीएल का मूल्यांकन, गैर-आक्रामक होने के दौरान, रिज़ॉल्यूशन का एक स्तर प्रदान करता है जो फाइबर के भीतर एलडी के आकार और सटीक स्थान का अध्ययन करने के लिए पर्याप्त नहीं है, और यह फाइबर-प्रकार विशिष्ट17,18 नहीं है। इसी तरह, पूरे मांसपेशी होमोजेनेट्स19 पर की गई जैव रासायनिक तकनीक लिपिड के स्थान और आकार का आकलन नहीं कर सकती है। नतीजतन, एलडी आकृति विज्ञान और स्थान का विश्लेषण करने का सबसे पर्याप्त तरीका मात्रात्मक संचरण इलेक्ट्रॉनिक माइक्रोस्कोपी13 है, लेकिन यह तकनीक महंगी और समय लेने वाली है। इसलिए, ऑयल रेड ओ (ओआरओ) 20,21, मोनोडैनसिलपेंटेन (एमडीएच) 22, या बोडिपी 23,24,25 जैसे रंगों के साथ तैयारी पर कॉन्फोकल फ्लोरेसेंस इमेजिंग, इन अध्ययनों के लिए सबसे अच्छा उपकरण के रूप में उभरा है।
यहां, माउस मांसपेशी क्रायोसेक्शन में एलडी आकार, संख्या और स्थानीयकरण को मापने के लिए ऊतक नमूनाकरण और प्रसंस्करण, बोडिपी धुंधला, और कन्फोकल छवि अधिग्रहण और विश्लेषण सहित एक पूर्ण प्रोटोकॉल का वर्णन किया गया है। चूंकि आईएमसीएल ऑक्सीडेटिव और ग्लाइकोलाइटिक फाइबर के बीच समान रूप से वितरित नहीं किए जाते हैं, और प्रत्येक फाइबर प्रकार एलडी गतिशीलता को अलग तरह से नियंत्रित करता है, आईएमसीएल का अध्ययन फाइबर-प्रकार विशिष्ट 16,25,26,27 होना चाहिए। इसलिए, यह प्रोटोकॉल प्रत्येक फाइबर द्वारा व्यक्त मायोसिन भारी श्रृंखला (एमवाईएचसी) आइसोफॉर्म (ओं) की पहचान करने के लिए धारावाहिक वर्गों पर इम्यूनोफ्लोरेसेंस का उपयोग करता है। इस प्रोटोकॉल का एक और लाभ एक ग्लाइकोलाइटिक (एक्सटेंसर डिजिटोरम लॉन्गस, ईडीएल) और एक ऑक्सीडेटिव (एकमात्र) मांसपेशियों के एक साथ प्रसंस्करण ठंड (चित्रा 1) से पहले कंधे से कंधा मिलाकर रखा गया है। यह एक साथ प्रसंस्करण न केवल समय बचाता है बल्कि नमूनों के अलग-अलग प्रसंस्करण के कारण परिवर्तनशीलता से भी बचता है।
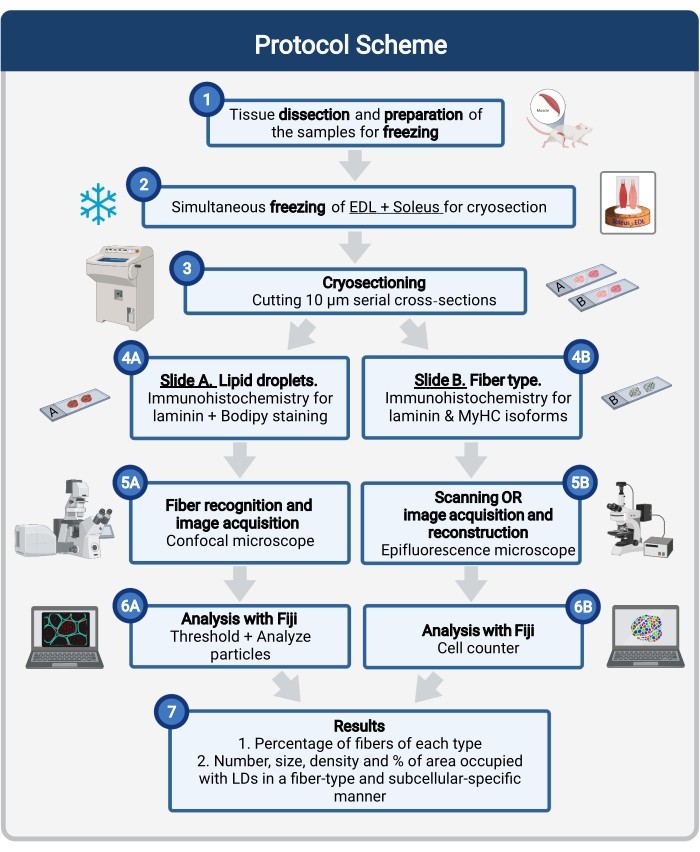
चित्रा 1: प्रक्रिया का योजनाबद्ध अवलोकन। मांसपेशियों के विच्छेदन (1) के बाद, समान आकार की चयनित मांसपेशियों को तैयार किया जाता है और एक साथ जमे हुए होते हैं (2). 10 μm के सीरियल अनुप्रस्थ वर्गों को क्रायोस्टैट का उपयोग करके प्राप्त किया जाता है और सीधे आसंजन स्लाइड (3) पर घुड़सवार किया जाता है। दो धारावाहिक स्लाइडों से, पहला (4 ए) लैमिनिन के लिए इम्यूनोलेबल किया जाता है और एलडी को पहचानने के लिए बोडिपी के साथ दाग दिया जाता है और दूसरा (4 बी) मांसपेशियों के फाइबर प्रकारों की पहचान के लिए माईएचसी के खिलाफ एंटीबॉडी के साथ इम्यूनोस्टेन किया जाता है। छवियों को बोडिपी (5 ए) के लिए एक कॉन्फोकल माइक्रोस्कोप और मांसपेशी फाइबर प्रकारों (5 बी) के लिए एक एपिफ्लोरेसेंस माइक्रोस्कोप का उपयोग करके अधिग्रहित किया जाता है। अनुभाग में प्रत्येक प्रकार के तंतुओं का प्रतिशत प्राप्त करने के लिए एलडी (7) या गिनती कोशिकाओं (6 बी) द्वारा कब्जा किए गए कुल क्षेत्र की संख्या, औसत आकार, घनत्व और प्रतिशत प्राप्त करने के लिए एक थ्रेसहोल्ड और मात्रा निर्धारित कणों (6 ए) को लागू करके फिजी में छवियों का विश्लेषण किया जाता है। संक्षिप्त नाम: एलडी = लिपिड बूंदें; ईडीएल = एक्सटेंसर डिजिटोरम लॉन्गस; मायएचसी = मायोसिन भारी श्रृंखला आइसोफॉर्म। कृपया इस आंकड़े का एक बड़ा संस्करण देखने के लिए यहां क्लिक करें।
