Strukturforskning i botanikk, som dekker plantemorfologi og anatomi, er grunnleggende for å forstå hele organismen1,2, og gir uunnværlige perspektiver for å integrere og bidra til kunnskap om økologi, fysiologi, utvikling og evolusjon av planter3. Metoder i plantemorfologi og anatomi omfatter i dag protokoller, utstyr og kunnskap utviklet nylig så vel som for mer enn et århundre siden2. Den kontinuerlige utførelsen og tilpasningen av klassiske metoder (f.eks. Lysmikroskopi) sammen med nyere teknikker (f.eks. Konfokalmikroskopi, røntgenmikrotomografi) har samme viktige grunnlag: teoretisk kunnskap som muliggjør utvikling av en metodikk.
Hovedverktøyet i planteanatomi og morfologi er bildet. Til tross for misforståelsen om at slike analyser er enkle observasjoner, gir plass til subjektive tolkninger2, krever analyse og forståelse av bilder på dette området kunnskap om metodene som brukes (utstyret, type analyse, metodologiske prosedyrer), cellekomponenter, histokjemi og plantekroppen (vevsorganisasjon og funksjon, ontogeni, morfologiske tilpasninger). Tolkning av bildene oppnådd ved hjelp av en rekke metoder kan føre til korrelerende form og funksjon, dechiffrere den kjemiske sammensetningen av en struktur, bekrefte i å beskrive taxa, forstå infeksjoner av fytopatogener og andre slike vurderinger.
Når man forsker på mykoheterotrofe (MH) planter (dvs. ikke-fotosyntetiske planter som får karbon fra mykorrhizal sopp4,5), kan bemerkelsesverdige aspekter av deres strukturelle tilpasninger, mønstrene for vevkolonisering av sopp og morfoanatomi av underjordiske organer opplyse deres utviklingsstrategier og forhold til hyfer, som er kilden til næringsstoffer. De underjordiske organene til MH-planter viser vanligvis viktige tilpasninger knyttet til deres tilknytning til jordsvamp, derfor er det viktig å utføre disse anatomiske og morfologiske undersøkelsene6. MH-arters luftorganer bør ikke ignoreres, da endofytter også kan være tilstede i disse vevene, selv om de ikke er mykorrhizale sopp (personlige observasjoner, ikke publisert ennå).
I tillegg til den veletablerte essensen av mykorrhizal soppforening med MH-arter i løpet av hele livssyklusen7, har hver orkidéart, selv de autotrofe, et innledende obligatorisk mykoheterotrofisk stadium i naturlige miljøer. Det oppstår fordi orkideenes embryo er utifferentiert og mangler endosperm eller cotyledoner, og dermed ikke er i stand til å utvikle og etablere seg i naturlige miljøer uten næringsstøtte fra sopppartnere 4,8. Tatt i betraktning at symbiotiske spiringsprotokoller kan brukes ikke bare på MH-arter, men også på fotosyntetiserende orkideer, med sikte på å undersøke orkidé-soppspesifisitet i spiring og protocormutvikling, en enormt anvendt metodikk i initiativer for bevaring av truede arter 9,10,11.
I denne metodesamlingen beskriver vi viktige trinn involvert i innsamling, fiksering og lagring av MH-planteprøver for anatomiske studier (seksjon 1), overflateanalyse og prøvevalg (seksjon 2), seksjoneringsmetoder (frihånd: seksjon 3, mikrotomi: seksjon 4, kryomikrotomi: seksjon 5), farging og montering (seksjon 6), fluorescens og konfokal mikroskopi av soppendofytter (seksjon 7), skanning elektronmikroskopi (seksjon 8), og transmisjonselektronmikroskopi (pkt. 9). I tillegg beskriver vi en symbiotisk spiringsmetode for orkidéfrø (MH og autotrofisk, seksjon 10), da de tidligere nevnte avbildningsmetodene med hell kan brukes til å analysere soppkolonisering av frø, protokormer og frøplanter i spiringsprosessen.
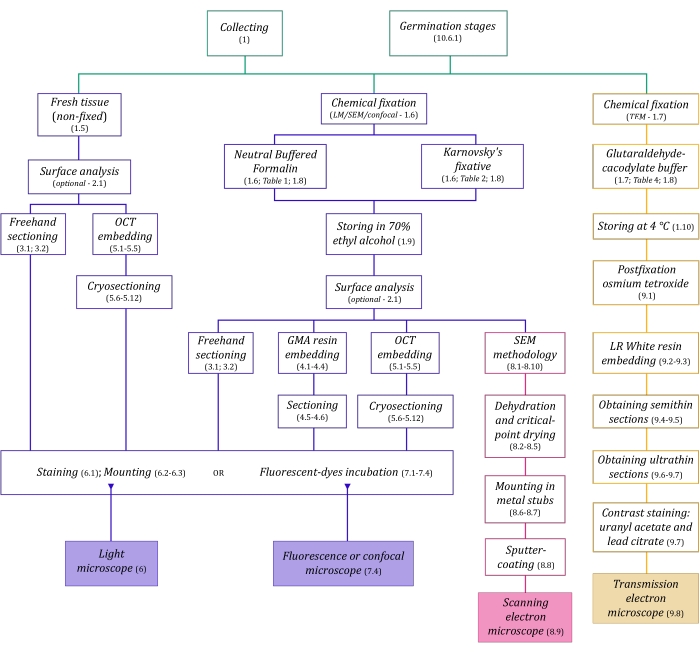
Figur 1: Skjematisk oppsummering av bildemetoder. Skjemaene gir indikasjoner på protokolltrinn der de er detaljerte. Forkortelser: GMA = glykolmetakrylat, OCT = optimal skjæretemperaturforbindelse, SEM = skanningelektronmikroskopi. Klikk her for å se en større versjon av dette tallet.
Mikroskopiteknikkene beskrevet her i detalj (figur 1) innledes av følgende viktige trinn: innsamling, festing, dehydrering, innebygging og seksjonering av prøver. Siden trinnene er variable (figur 1) avhengig av valgt(e) teknikk(er), er det viktig å tenke fremover, med tanke på fikseringsmidlene som skal klargjøres og transporteres til innsamlingsstedet, hvordan prøvene må klargjøres før fiksering, dehydreringsprosessene som skal brukes (avsnitt 1), og ulike innbyggingsmuligheter og snittingsmetoder (§§ 4, 5, og 9). Figur 1 oppsummerer sekvensielt alle trinnene som kreves for hver mikroskopiteknikk grundig beskrevet nedenfor.
