בגוף האדם חיים כ-10-100 טריליון תאים מיקרוביאליים חיים (חיידקים, פטריות ארכאיות), המצויים בעיקר בסביבות המעיים, העור והרירית1. במצב בריא, אלה מספקים יתרונות למארח שלהם, כולל ייצור ויטמינים, התבגרות של מערכת החיסון, גירוי של תגובות חיסוניות מולדות ונרכשות לפתוגנים, ויסות חילוף החומרים של השומן, ויסות תגובות ללחץ ועוד, עם השפעה על גדילה והתפתחות, התפרצות מחלות והזדקנות 2,3,4,5 . גם המיקרוביוטה של המעיים מתפתחת במידה ניכרת במהלך החיים. האבולוציה הדרסטית ביותר מתרחשת במהלך הינקות והילדות המוקדמת6, אך שינויים משמעותיים מתרחשים גם עם הגיל, כולל ירידה בשפע ביפידובקטריום ועלייה במינים קלוסטרידיום, לקטובצילוס, אנטרובקטריה ואנטרוקוקוס 7. אורח חיים יכול לשנות עוד יותר את ההרכב המיקרוביאלי של המעיים ולהוביל לדיסביוזיס (אובדן חיידקים מועילים, צמיחת יתר של חיידקים אופורטוניסטיים), וכתוצאה מכך פתולוגיות שונות כגון מחלות מעי דלקתיות, סוכרת והשמנת יתר5, אך גם לתרום למחלות אלצהיימר ופרקינסון 8,9,10,11.
הבנה זו תרמה תרומה קריטית לעידון הרעיון של ציר המעי-מוח (GBA), שבו אינטראקציות בין הפיזיולוגיה של המעיים (כיום כולל המיקרובים שבתוכה) לבין מערכת העצבים נחשבות לווסת העיקרי של חילוף החומרים של בעלי החיים ולתפקודים פיזיולוגיים12. עם זאת, התפקיד המדויק של המיקרוביוטה באיתות בין המעי למוח ומנגנוני הפעולה הקשורים אליו רחוקים מלהיות מובנים במלואם13. מאחר שמיקרוביוטה של המעיים היא גורם מפתח להזדקנות בריאה, האופן שבו חיידקים מווסתים את תהליך ההזדקנות הפך לנושא למחקר אינטנסיבי ולמחלוקת 6,14,15.
עם ההדגמה שהתולעת העגולה Caenorhabditis elegans מארחת מיקרוביוטה של המעי בונפיד הנשלטת – כמו במינים אחרים – על ידי Bacteroidetes, Firmicutes ו- Actinobacteria 16,17,18,19,20, עלייתה המהירה כפלטפורמה ניסיונית לחקר אינטראקציות בין המעי המארח למעיים 21,22,23,24 ,25,26 הרחיב משמעותית את ארסנל החקירות שלנו26,27,28,29. בפרט, ניתן להתאים גישות ניסיוניות בעלות תפוקה גבוהה הזמינות עבור C. elegans לחקר תזונה גנטית, גנים-תרופות, גנים-פתוגנים וכו’, כדי לחקור במהירות כיצד חיידקים מבודדים וקוקטיילים משפיעים על הבריאות וההזדקנות של C. elegans.
הפרוטוקול הנוכחי מתאר צינור ניסיוני לסינון בבת אחת של מערכים של חיידקים מבודדים או תערובות שנקבעו בלוחות מרובי בתים להשפעות על עמידות ללחץ C. elegans כפרוקסי לבריאות, אשר ניתן להשתמש בהם כדי לזהות פרוביוטיקה. הוא מפרט כיצד לגדל אוכלוסיות תולעים גדולות ולטפל במערכי חיידקים בתבניות של לוחות 96 ו-384 בארות לפני עיבוד תולעים לניתוח אוטומטי של עמידות בפני עקה באמצעות קורא לוחות פלואורסצנטיים (איור 1). הגישה מבוססת על מבחני הישרדות אוטומטיים ללא תוויות (LFASS)30 המנצלים את תופעת פלואורסצנציית המוות31, לפיה תולעים גוססות מייצרות פרץ של פלואורסצנציה כחולה שניתן להשתמש בה כדי לאתר את זמן המוות. פלואורסצנציה כחולה נפלטת על ידי אסטרים של גלוקוזיל של חומצה אנתרנילית המאוחסנים בגרגרי מעיים של C. elegans (סוג של אברונים הקשורים לליזוזום), אשר מתפוצצים כאשר מפל נמק מופעל במעי התולעת עם המוות31.
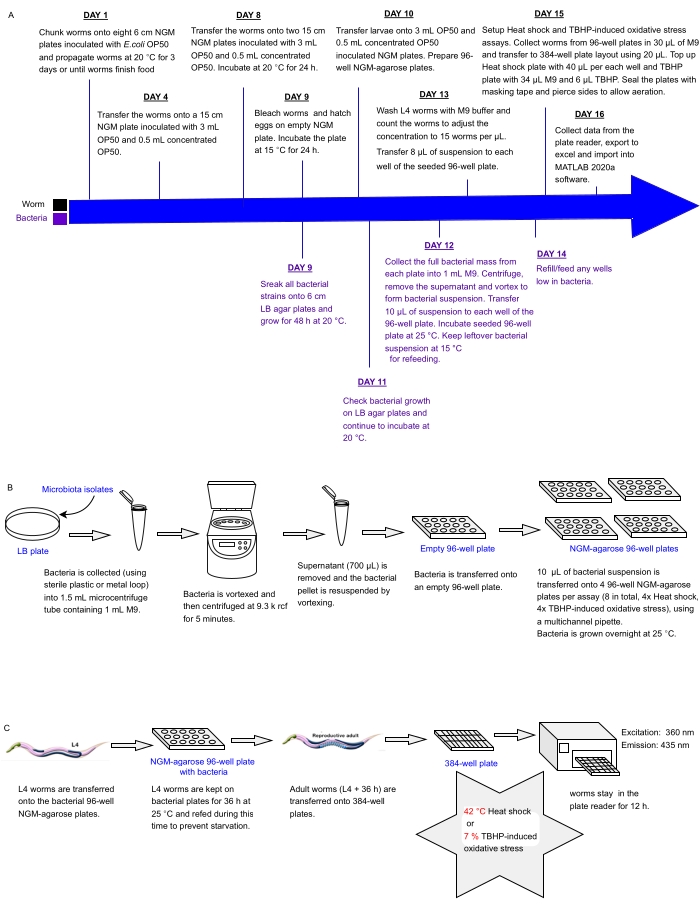
איור 1: זרימת עבודה ניסיונית לסינון בתפוקה גבוהה של חיידקים מבודדים עם השפעה על עמידות C. elegans ללחץ . (A) ציר זמן לתחזוקה והגדרת בדיקה של תולעים וחיידקים. (B) מערך לוחות חיידקים 96 באר התקנה וטיפול. (C) 384-באר צלחת תולעת. אנא לחץ כאן כדי להציג גרסה גדולה יותר של נתון זה.
