يتوفر عدد لا يحصى من النماذج لدراسة تطور السرطان ، كل منها فريد من نوعه ويمثل نوعا فرعيا من هذا المرض المعقد. يوفر كل نموذج رؤى فريدة وقيمة في بيولوجيا السرطان وقد حسن الوسائل لمحاكاة حالة المرض الفعلية. قدمت خطوط الخلايا الراسخة التي نمت كطبقة أحادية رؤى قيمة في العمليات الحيوية في المختبر ، مثل الانتشار والغزو والهجرة وموت الخلايا المبرمج1. على الرغم من أن زراعة الخلايا ثنائية الأبعاد (2D) كانت الأداة التقليدية للتحقيق في استجابة خلايا الثدييات للعديد من الاضطرابات البيئية ، إلا أن استقراء هذه النتائج للتنبؤ بالاستجابات على مستوى الأنسجة لا يبدو مقنعا بما فيه الكفاية. القيد الرئيسي للثقافات ثنائية الأبعاد هو أن البيئة الدقيقة التي تم إنشاؤها تختلف إلى حد كبير عن تلك الموجودة في أنسجة الثدي نفسها2. تفتقر ثقافة 2D إلى تفاعل الخلايا مع المصفوفة خارج الخلية ، وهو أمر حيوي لنمو أي أنسجة. أيضا ، فإن قوى الشد التي تعاني منها الخلية في مزارع الطبقة الأحادية تعيق قطبية هذه الخلايا ، وبالتالي تغير إشارات الخلية وسلوكها3،4،5. فتحت أنظمة الاستزراع ثلاثية الأبعاد (3D) طريقا جديدا في مجال أبحاث السرطان مع قدرتها على محاكاة الظروف في الجسم الحي في المختبر. يمكن إعادة إنشاء العديد من الإشارات البيئية الدقيقة الحاسمة التي تضيع في زراعة الخلايا ثنائية الأبعاد باستخدام ثقافات ثلاثية الأبعاد من المصفوفة خارج الخلية الغنية باللامينين (lrECM)6.
حددت دراسات مختلفة أهمية البيئة الدقيقة للورم في التسرطن 7,8. العوامل المرتبطة بالالتهاب هي جزء رئيسي من البيئة الدقيقة. عامل تنشيط الصفائح الدموية (PAF) هو وسيط فوسفوليبيد تفرزه خلايا مناعية مختلفة تتوسط استجابات مناعية متعددة 9,10. يتم إفراز مستويات عالية من PAF بواسطة خطوط خلايا سرطان الثدي المختلفة وترتبط بالانتشار المعزز11. أظهرت الدراسات التي أجريت في مختبرنا أن الوجود المطول ل PAF في الثقافات الأسينارية يؤدي إلى تحول الخلايا الظهارية للثدي12. يقوم PAF بتنشيط مستقبل PAF (PAFR) ، وتنشيط محور الإشارة PI3K / Akt13. وتفيد التقارير أيضا أن PAFR مرتبط ب EMT والغزو والانبثاث14.
يوضح هذا البروتوكول نظاما نموذجيا لدراسة التحول الناجم عن PAF ، باستخدام ثقافات ثلاثية الأبعاد للخلايا الظهارية للثدي ، كما وصفها سابقا Chakravarty et al.12. تميل الخلايا الظهارية للثدي التي تنمو على المصفوفة خارج الخلية (ثقافات 3D) إلى تشكيل كرويات مستقطبة توقف النمو. وتسمى هذه الأسيني وتشبه إلى حد كبير الأسيني من أنسجة الثدي ، أصغر وحدة وظيفية في الغدة الثديية ، في الجسم الحي15. تتكون هذه الكرويات (الشكل 1A ، B) من طبقة واحدة من الخلايا الظهارية المستقطبة المعبأة بشكل وثيق حول تجويف مجوف ومتصلة بالغشاء السفلي (الشكل 1C). تم وصف عملية التشكل هذه بشكل جيد في الأدب16. عند زرعها على lrECM ، تخضع الخلايا للانقسام والتمايز لتشكيل مجموعة من الخلايا ، والتي تستقطب بعد ذلك من اليوم 4 فصاعدا. بحلول اليوم 8 ، يتكون الأسيني من مجموعة من الخلايا المستقطبة التي تكون على اتصال مباشر مع المصفوفة خارج الخلية ومجموعة من الخلايا غير المستقطبة المغلقة داخل الخلايا المستقطبة الخارجية ، مع عدم وجود اتصال بالمصفوفة . من المعروف أن هذه الخلايا غير المستقطبة تخضع لموت الخلايا المبرمج بحلول اليوم 12 من الثقافة ، مما يشكل تجويفا مجوفا. بحلول اليوم 16 ، يتم تشكيل الهياكل المتوقفة عن النمو16.
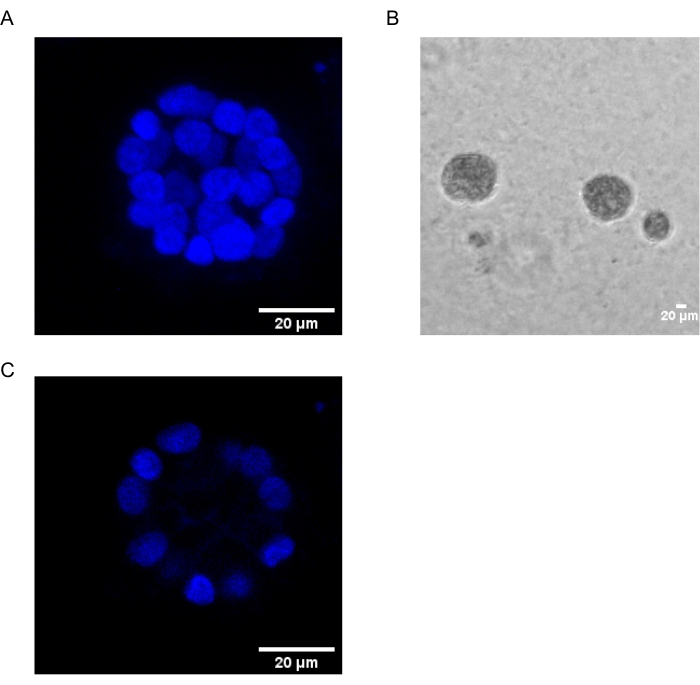
الشكل 1: نوى الخلايا في الأسيني الملطخة ببقعة نووية . (A) بناء 3D من الأسيني. (ب) صورة تباين الطور ل MCF10A acini المزروعة في Matrigel لمدة 20 يوما. (ج) يظهر القسم الأوسط وجود تجويف مجوف. شريط المقياس = 20 ميكرومتر. يرجى النقر هنا لعرض نسخة أكبر من هذا الرقم.
على عكس ثقافات 2D ، تساعد ثقافات acinar في تمييز الخلايا الطبيعية والمحولة من خلال التغيرات المورفولوجية الواضحة. تشكل الخلايا الظهارية غير المتحولة في الثدي الأسيني مع تجويف مجوف ، يحاكي أسيني الثدي البشري الطبيعي. تظهر هذه الكرويات ، عند التحول ، مورفولوجيا معطلة تتميز بفقدان كبير للقطبية (واحدة من السمات المميزة للسرطان) ، أو عدم وجود تجويف ، أو اضطراب في التجويف المجوف (بسبب التهرب من موت الخلايا المبرمج) الذي قد يحدث بسبب تحرير الجينات المختلفة17،18،19،20 . يمكن دراسة هذه التحولات باستخدام تقنيات شائعة الاستخدام مثل التألق المناعي. وبالتالي ، يمكن أن يعمل نموذج زراعة الخلايا 3D كطريقة بسيطة للتحقيق في عملية تكوين الثدي وسرطان الثدي. إن إنشاء نظام ثقافة 3D لفهم تأثير وسيط الدهون الفوسفاتية ، PAF ، سيساعد في فحص الأدوية قبل السريرية عالي الإنتاجية.
قام هذا العمل بتكييف بروتوكول الثقافة ثلاثية الأبعاد “على القمة”16,21 لدراسة التحول الناجم عن PAF 22. تمت دراسة التغيرات المظهرية الناجمة عن تعرض الأسيني لوسيط الدهون الفوسفاتية باستخدام التألق المناعي. تم استخدام العديد من علامات القطبية والظهارية إلى الانتقال الوسيط (EMT) 12,16 في الدراسة. ويذكر الجدول 1 توطينها الطبيعي ونمطها الظاهري المتوقع عند التحول.
| الاجسام المضاده | علامات | التوطين العادي | النمط الظاهري المحول |
| α6-إنتيغرين | القاعدية | القاعدية مع وصمة عار جانبية ضعيفة | وصمة عار جانبية / قمية قوية |
| β كاتينين | تقاطع الخلية والخلية | القاعدية | توطين غير طبيعي / نووي أو سيتوبلازمي |
| فيمنتين | EMT | غياب / ضعف الحضور | التنظيم المرتفع |
الجدول 1: العلامات المستخدمة في الدراسة. علامات مختلفة تستخدم مع توطينها في وجود وغياب علاج PAF.
يمكن استخدام هذه الطريقة بشكل أفضل لدراسة / فحص الأدوية المعقولة والجينات المستهدفة لمختلف الأنواع الفرعية لسرطان الثدي. وهذا يمكن أن يوفر بيانات استجابة للأدوية أقرب إلى سيناريو في الجسم الحي ، مما يساعد في تطوير الأدوية بشكل أسرع وأكثر موثوقية. أيضا ، يمكن استخدام هذا النظام لدراسة الإشارات الجزيئية المرتبطة بالاستجابة للأدوية ومقاومة الأدوية.
