Kanserin ilerlemesini incelemek için sayısız model mevcuttur, bunların her biri benzersizdir ve bu karmaşık hastalığın bir alt tipini temsil etmektedir. Her model, kanser biyolojisine benzersiz ve değerli bilgiler sağlar ve gerçek hastalık durumunu taklit etme araçlarını geliştirmiştir. Tek katmanlı olarak yetiştirilen yerleşik hücre hatları, proliferasyon, invazivlik, göç ve apoptoz1 gibi in vitro hayati süreçler hakkında değerli bilgiler sağlamıştır. İki boyutlu (2D) hücre kültürü, memeli hücrelerinin çeşitli çevresel karışıklıklara tepkisini araştırmak için geleneksel bir araç olmasına rağmen, doku düzeyindeki yanıtları tahmin etmek için bu bulguların ekstrapolasyonu yeterince ikna edici görünmemektedir. 2D kültürlerin en büyük sınırlaması, yaratılan mikro çevrenin meme dokusunun kendisinden büyük ölçüde farklı olmasıdır2. 2D kültür, hücrelerin herhangi bir dokunun büyümesi için hayati önem taşıyan hücre dışı matris ile etkileşiminden yoksundur. Ayrıca, tek katmanlı kültürlerde hücrenin yaşadığı gerilme kuvvetleri, bu hücrelerin polaritesini engeller, böylece hücre sinyalini ve davranışını değiştirir 3,4,5. Üç boyutlu (3D) kültür sistemleri, in vivo koşulları in vitro olarak taklit etme yetenekleriyle kanser araştırmaları alanında yeni bir yol açmıştır. 2B hücre kültüründe kaybolan birçok önemli mikroçevresel ipucu, laminin bakımından zengin hücre dışı matrisin (lrECM)6 3B kültürleri kullanılarak yeniden oluşturulabilir.
Çeşitli çalışmalar karsinogenezde tümör mikroçevresinin önemini belirlemiştir 7,8. Enflamasyonla ilişkili faktörler mikro çevrenin önemli bir parçasıdır. Trombosit Aktive Edici Faktör (PAF), çeşitli immün hücreler tarafından salgılanan ve çoklu immün yanıtlara aracılık eden bir fosfolipid mediatörüdür 9,10. Yüksek PAF seviyeleri farklı meme kanseri hücre hatları tarafından salgılanır ve artmış proliferasyon ile ilişkilidir11. Laboratuvarımızdan yapılan çalışmalar, asiner kültürlerde PAF’ın uzun süreli varlığının meme epitel hücrelerinin transformasyonuna yol açtığını göstermiştir12. PAF, PAF reseptörünü (PAFR) aktive ederek PI3K/Akt sinyal ekseni13’ü aktive eder. PAFR’nin ayrıca EMT, invazyon ve metastaz14 ile ilişkili olduğu bildirilmiştir.
Mevcut protokol, daha önce Chakravarty ve ark.12 tarafından tanımlandığı gibi, meme epitel hücrelerinin 3D kültürlerini kullanarak, PAF kaynaklı transformasyonu incelemek için bir model sistemi göstermektedir. Hücre dışı matriste (3D kültürler) yetiştirilen meme epitel hücreleri, polarize büyüme ile tutuklanan sferoidler oluşturma eğilimindedir. Bunlara acini denir ve meme bezinin en küçük fonksiyonel birimi olan meme dokusunun akinisine in vivo 15’e çok benzer. Bu sferoidler (Şekil 1A,B), içi boş bir lümeni çevreleyen ve bazal membrana tutturulmuş, sıkıca paketlenmiş polarize epitel hücrelerinden oluşan bir tek katmandan oluşur (Şekil 1C). Bu morfogenez süreci literatür16’da iyi tanımlanmıştır. LrECM üzerine tohumlandığında, hücreler bir hücre kümesi oluşturmak için bölünme ve farklılaşmaya uğrar ve daha sonra 4. Gün’den itibaren polarize olurlar. 8. Günde, acini, hücre dışı matrisle doğrudan temas halinde olan bir grup polarize hücreden ve dış polarize hücrelerin içine kapatılmış, matrise temas etmeyen polarize edilmemiş bir hücre kümesinden oluşur. Bu polarize olmayan hücrelerin, kültürün 12. Gününe kadar apoptoza uğradığı ve içi boş bir lümen oluşturduğu bilinmektedir. 16. Gün’e gelindiğinde, büyümeyi durduran yapılar16.
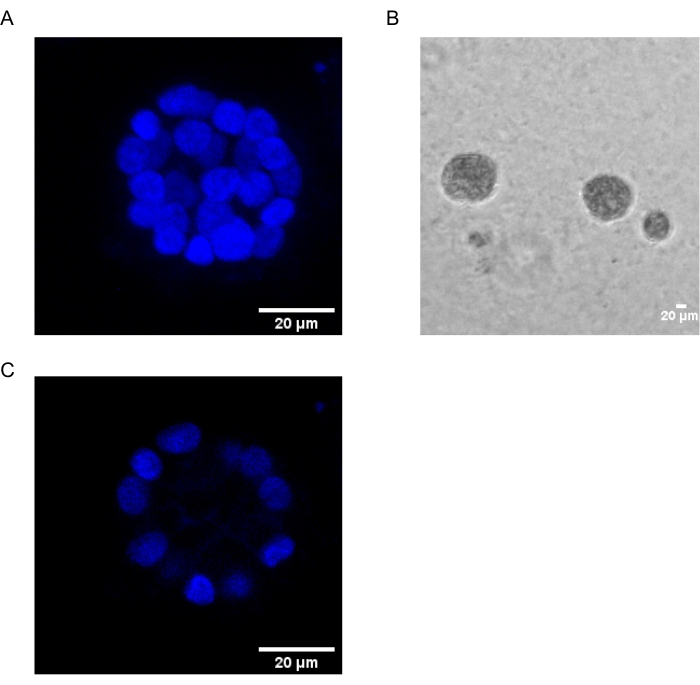
Şekil 1: Acini’deki hücrelerin çekirdekleri nükleer bir leke ile boyanmıştır . (A) Acini’nin 3D yapısı. (B) Matrigel’de 20 gün boyunca yetiştirilen MCF10A acini’nin Faz Kontrastı görüntüsü. (C) En ortadaki bölüm içi boş bir lümenin varlığını gösterir. Ölçek çubuğu = 20 μm. Bu şeklin daha büyük bir versiyonunu görüntülemek için lütfen buraya tıklayın.
2D kültürlerin aksine, acinar kültürler normal ve dönüştürülmüş hücreleri görünür morfoloji değişiklikleri yoluyla ayırt etmeye yardımcı olur. Transforme olmayan meme epitel hücreleri, normal insan meme acinisini taklit eden içi boş bir lümen ile acini oluşturur. Bu sferoidler, transformasyon üzerine, çeşitli genlerin deregülasyonuna bağlı olarak indüklenebilecek büyük bir polarite kaybı (kanserin ayırt edici özelliklerinden biri), bir lümen yokluğu veya içi boş lümenin bozulması (apoptozun kaçması nedeniyle) ile karakterize bozulmuş bir morfoloji göstermektedir17,18,19,20 . Bu dönüşümler, immünofloresan gibi yaygın olarak kullanılan teknikler kullanılarak incelenebilir. Böylece, 3D hücre kültürü modeli, meme asinar morfogenezi ve meme kanserogenezi sürecini araştırmak için basit bir yöntem olarak işlev görebilir. Bir fosfolipit mediyatörü olan PAF’ın etkisini anlamak için bir 3D kültür sistemi kurmak, yüksek verimli preklinik ilaç taramasına yardımcı olacaktır.
Bu çalışma, PAF 22 tarafından indüklenen dönüşümü incelemek için 3D ‘üstte’ kültür protokolü 16,21’i uyarladı. Acının fosfolipid mediatöre maruz kalmasıyla indüklenen fenotipik değişiklikler immünofloresan kullanılarak incelendi. Çalışmada çeşitli polarite ve epitelyal-mezenkimal geçiş (EMT) belirteçleri 12,16 kullanıldı. Tablo 1’de normal lokalizasyonlarından ve transformasyon üzerine beklenen fenotiplerinden bahsedilmektedir.
| Antikor | Işaret | Normal lokalizasyon | Dönüştürülmüş fenotip |
| α6-İntegrin | Basolateral | Zayıf yanal lekeli bazal | Güçlü yanal / apikal leke |
| β-Katenin | Hücre-hücre bağlantısı | Basolateral | Anormal / nükleer veya sitoplazmik lokalizasyon |
| Arjantin | Emt | Eksik / zayıf mevcudiyet | Yukarı düzenleme |
Tablo 1: Çalışmada kullanılan belirteçler. PAF tedavisinin varlığında ve yokluğunda lokalizasyonları ile kullanılan farklı belirteçler.
Bu yöntem, makul ilaçları incelemek / taramak ve çeşitli meme kanseri alt tipleri için genleri hedeflemek için en iyi şekilde kullanılabilir. Bu, in vivo senaryoya daha yakın bir ilaç yanıt verisi sağlayarak daha hızlı ve daha güvenilir ilaç geliştirilmesine yardımcı olabilir. Ayrıca, bu sistem ilaç yanıtı ve ilaç direnci ile ilişkili moleküler sinyalizasyonu incelemek için kullanılabilir.
