In this article, a detailed protocol for the isolation of conditionally immortalized glomerular endothelial cells with stable fluorescent mitochondria (mitoDendra2-GECs) is described (Figure 1). The use of young 6-10-week-old mice is essential to obtain a substantial number of healthy cells. After 3 days of culture, cells start growing slowly from the isolated glomeruli, as shown in Figure 3G. After 7 days, cells are heterogeneous, showing other glomerular cell types, such as podocytes, parietal epithelial cells, and mesangial cells (Figure 4A). After reaching 70% to 80% confluency, other glomerular cells were removed using a positive selection of endothelial cells with magnetic CD31 labeled beads (Figure 4B–C). The mitoDendra2-GECs expressed CD31 (Figure 4D) and were negative for podocyte marker as shown by synaptopodin negative staining (Figure 4E-F).
The fluorescent mitochondrial feature in the mitoDendra2-GECs allows for studying mitochondria morphology under various conditions in vitro. Here, we tested the effect of high glucose (25mM) on the mitochondria structure of mitoDendra2-GECs (Figure 5D-F). Compared to the elongated mitochondria visible in cells under normal glucose (5mM; Figure 5A-C), high glucose-induced fragmentation or fission of mitochondria as observed by prominent spheroid-shaped mitochondria (Figure 5E). Furthermore, we used a 405 nm laser to photo-convert a selected subpopulation of mitochondria in a single live mitoDendra2-GEC (Figure 5A,D). In Figure 5B and Figure 5E, successful photo-switching of mitochondria from green (488nm) to red (561 nm) in the selected area is presented. Importantly, it allowed for witnessing the fusion events in mitoDendra2-GECs grown under normal glucose, as can be observed by the yellow merged green and red fluorescence as a result of mitochondria matrix fusion (Figure 5C). In contrast, in the high glucose-treated mitoDendra2-GEC, the mitochondria were fragmented and were mainly red (Figure 5F), suggesting that, in the time frame tested, the fission/fusion events were delayed or inhibited due to the damage caused by the high glucose treatment, and this is consistent with previous reports18,19.
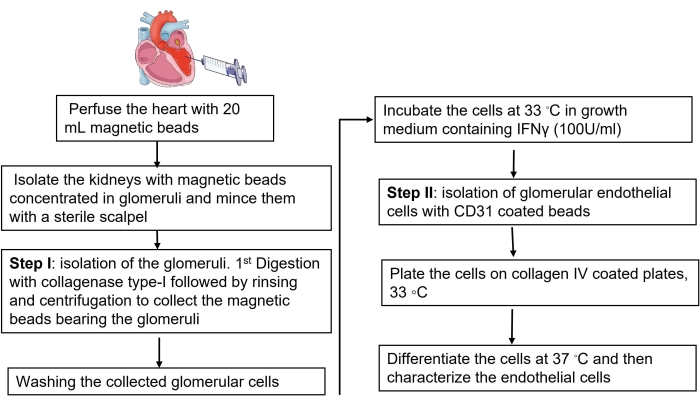
Figure 1: Representative description of murine GEC isolation. The diagram represents the significant steps of GEC isolation, starting from perfusing mice heart with magnetic beads to isolate the kidney glomeruli, the digestion of collagenase, the culture of glomerular cells, and finally, the purification of GEC using CD31-coated beads. Please click here to view a larger version of this figure.
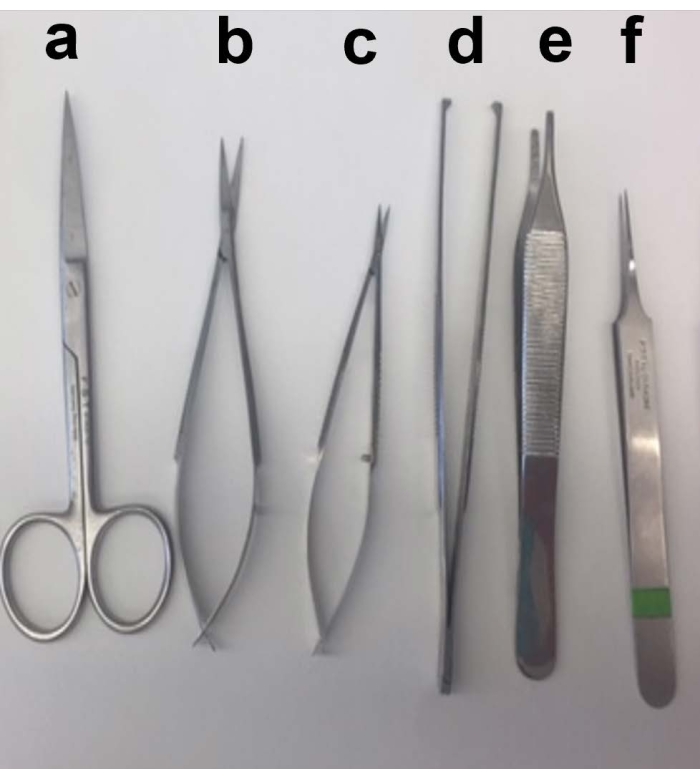
Figure 2: Description of kidney dissection. Representative image of all the sterile surgical instruments needed for the dissection. Scissors (a) are needed to open the skin of the mouse, and a separate pair of scissors (b,c) are needed to isolate the kidney. Tweezers (d) and (e) are needed to hold the skin. Tweezer (f) is needed to collect the dissected kidney. Please click here to view a larger version of this figure.
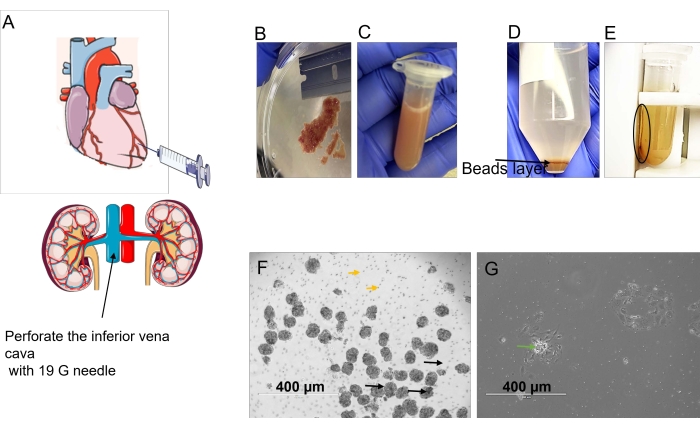
Figure 3: Description of glomeruli harvesting. (A) Perfuse the left ventricle with the HBSS-beads solution using a 20 mL syringe. Perforate the inferior vena cava with a 19 G needle. (B) After perfusion, remove the fat from the kidney and cut it into 1 mm pieces. (C) Digest the tissue with collagenase type II. (D) After digestion and centrifugation, three layers are formed; the beads bearing the glomeruli are in the middle layer, as indicated by the black arrow. (E) Separate the isolated beads bearing the glomeruli using the magnetic concentrator. (F) Representative image of freshly isolated glomeruli. (G) Growing cells from isolated glomeruli after 3 days. Please click here to view a larger version of this figure.
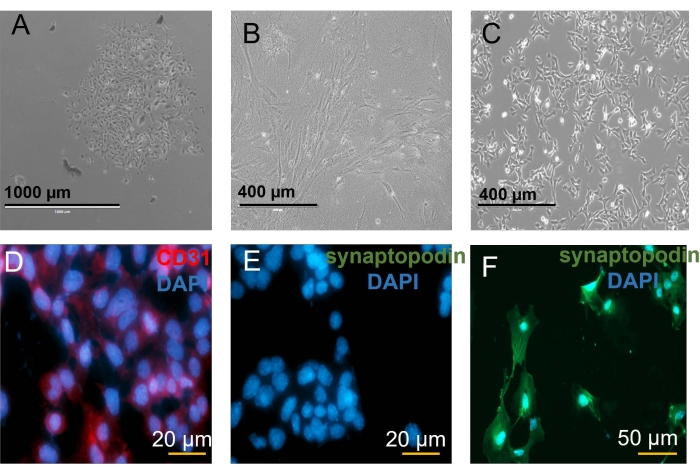
Figure 4: Mitochondrial assay in vitro. (A) Glomerular cells after 3 days of culture. (B) GEC after 7 days. (D) Purification of GEC with CD31-labeled beads. Microscopic imaging of CD31 staining. (E) GECs are negative for synaptopodin. (F) Podocytes stained with synaptopodin (488 nm, green fluorescence) as a negative control. Please click here to view a larger version of this figure.
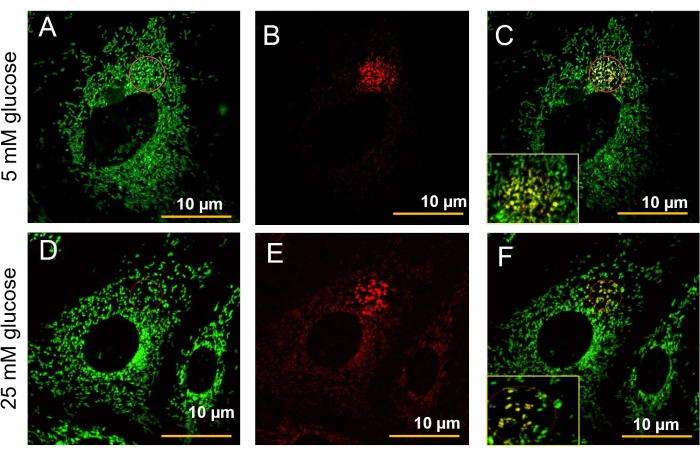
Figure 5: Microscopic imaging of fluorescent mitochondrial fusion in vitro. Images were acquired with a confocal microscope using a 40x water objective. (A–B) Selected area (red circle) was illuminated with a 405 nm line (4% laser power) for 300 bleaching iterations at a speed of 6.3-12.61/pixel, as previously described16. (A–C) Cells in growth media containing 5 mM glucose; green fluorescence (488 nm) shows healthy mitochondria. (B,E) Mitochondria were photo-switched into red (567 nm). (C) Yellow fluorescence detects an active fusion event of GEC treated with 5mM glucose. (D) Cells were treated with high glucose 25 mM for 30 min. Green fluorescence shows fragmented mitochondria and (E) a portion that was photo-switched into red. (F) The figure shows a decreased fusion event as shown by reduced yellow fluorescence. Please click here to view a larger version of this figure.
