Senescente celler akkumuleres normalt i organismer over år under normal biologisk aldring, men kan også udvikle sig hurtigt i tumorceller som et svar på skader induceret af forskellige kræftbehandlinger, herunder stråling og kemoterapi. Selvom de ikke længere formerer sig, kan terapiinducerede senescent (TIS) tumorceller bidrage til behandlingsresistens og drive gentagelse 1,2,3. Faktorer udskilt af TIS-celler kan forværre tumormalignitet ved at fremme immununddragelse eller metastase 4,5. TIS-celler udvikler komplekse, kontekstspecifikke fænotyper, ændrede metaboliske profiler og unikke immunresponser 6,7,8. Derfor er identifikation og karakterisering af TIS tumorceller induceret af forskellige kræftbehandlingsmetoder et emne af løbende interesse for kræftforskningssamfundet.
Til påvisning af TIS-tumorceller anvendes konventionelle ældningsassays i vid udstrækning, primært baseret på påvisning af øget aktivitet af ældningsmarkørenzymet, den lysosomale beta-galactosidase GLB19. Detektion ved en næsten neutral (snarere end sur) lysosomal pH muliggør specifik påvisning af ældningsassocieret beta-galactosidase (SA-β-Gal)10. Et standard SA-β-Gal-assay, der er blevet brugt i flere årtier, bruger X-Gal (5-brom-4-chlor-3-indolyl-β-D-galactopyranosid), et blåt kromogent beta-galactosidasesubstrat, til at detektere SA-β-Gal i faste celler ved lysmikroskopi11. X-Gal-analysen muliggør kvalitativ visuel bekræftelse af TIS ved hjælp af almindeligt tilgængelige reagenser og laboratorieudstyr. Et grundlæggende transmitteret lysmikroskop er den eneste instrumentering, der kræves for at evaluere tilstedeværelsen af det blå kromogen. X-Gal-farvningsproceduren kan dog mangle følsomhed, hvilket nogle gange kræver mere end 24 timer, før farven udvikler sig. Farvning efterfølges af subjektiv scoring med lav gennemstrømning af individuelle senescente celler baseret på tælling af cellerne, der udviser en vis intensitet af det blå kromogen under et lysmikroskop. Da X-Gal er celle-uigennemtrængelig, kræver dette assay opløsningsmiddelfikserede celler, som ikke kan genvindes til downstream-analyse. Når du arbejder med begrænsede prøver fra dyr eller patienter, kan dette være en stor ulempe.
Forbedrede SA-β-Gal-assays ved anvendelse af cellepermeant, fluorescerende enzymsubstrater, herunder C 12-FDG (5-dodecanoylaminofluorescein Di-β-D-Galactopyranosid, grøn) og DDAOG (9H-(1,3-dichlor-9,9-dimethylacridin-2-en-7-yl) β-D-Galactopyranosid, far-red) har tidligere optrådt i litteraturen12,13,14,15. DDAOG’s kemiske sondestruktur og optiske egenskaber er vist i supplerende figur S1. Disse cellepermeanerede sonder tillader analyse af levende (snarere end faste) celler, og fluorescerende snarere end kromogene sonder letter brugen af hurtige fluorescerende analyseplatforme med høj kapacitet, herunder screeningsinstrumenter med højt indhold og flowcytometre. Sortering af flowcytometre muliggør genopretning af berigede populationer af levende senescente celler fra cellekulturer eller tumorer til downstream-analyse (f.eks. Western blotting, ELISA eller ‘omics). Fluorescensanalyse giver også et kvantitativt signal, der giver mulighed for mere nøjagtig bestemmelse af procentdelen af senescente celler i en given prøve. Yderligere fluorescerende sonder, herunder levedygtighedssonder og fluorophoremærkede antistoffer, kan let tilføjes til multipleksanalyse af mål ud over SA-β-Gal.
I lighed med DDAOG er C12-FDG en fluorescerende sonde til SA-β-Gal, men dens grønne fluorescerende emission overlapper med iboende cellulær AF, som opstår under ældning på grund af akkumulering af lipofuscinaggregater i celler16. Ved at bruge den langt røde DDAOG-sonde kan grøn cellulær AF bruges som en sekundær parameter til at bekræfte ældning17. Dette forbedrer analysepålideligheden ved at bruge en anden markør ud over SA-β-Gal, som ofte kan være upålidelig som en enkelt markør for ældning18. Da påvisning af endogen AF i senescente celler er en etiketfri tilgang, er det en hurtig og enkel måde at udvide specificiteten af vores DDAOG-baserede assay.
I denne protokol demonstrerer vi brugen af DDAOG og AF som et hurtigt flowcytometriassay med to parametre til identifikation af levedygtige TIS-tumorceller fra in vitro-kulturer eller isoleret fra lægemiddelbehandlede tumorer etableret hos mus (figur 1). Protokollen bruger fluorophorer, der er kompatible med en bred vifte af standard kommercielle flowcytometrianalysatorer og sorterere (tabel 1). Kvantificering af procentdelen af levedygtige senescentceller ved hjælp af standard flowcytometrianalyse er aktiveret. Hvis det ønskes, kan der udføres et valgfrit immunmærkningstrin for at evaluere celleoverfladeantigener af interesse samtidig med ældning. Identificerede senescente celler kan også beriges ved hjælp af standard fluorescensaktiveret cellesorteringsmetode (FACS).
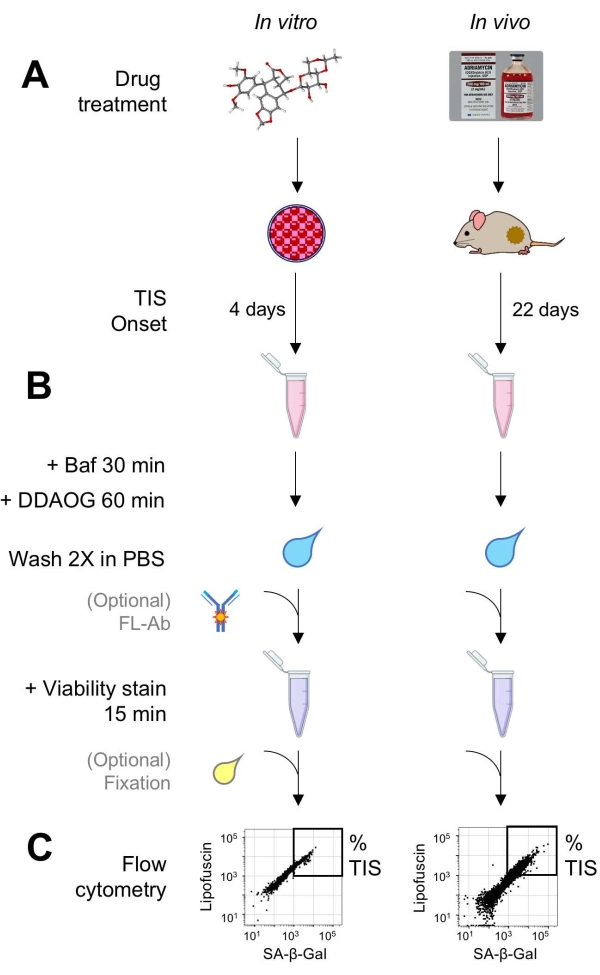
Figur 1: Eksperimentel arbejdsgang. En skematisk opsummering af nøglepunkter i DDAOG-analysen. (A) Et TIS-inducerende lægemiddel tilsættes til pattedyrsdyrkede celler eller administreres til tumorbærende mus. Der er derefter tid til at begynde med TIS: for celler, 4 dage efter behandling; for mus, 22 dage i alt, med tre behandlinger hver 5. dag plus 7 dages restitution. Celler høstes, eller tumorer dissocieres i suspension. (B) Prøver behandles med Baf for at justere lysosomal pH til påvisning af SA-β-Gal i 30 min. derefter tilsættes DDAOG-sonde i 60 minutter for at detektere SA-β-Gal. Prøver vaskes 2x i PBS, og der tilsættes kort en levedygtighedsplet (15 min). Eventuelt kan prøver farves med fluorescerende antistoffer i åbne fluorescenskanaler og / eller fastgøres til senere analyse. (C) Prøver analyseres ved hjælp af et standard flowcytometer. Levedygtige celler visualiseres i prikplotter, der viser rød DDAOG (indikerer SA-β-Gal) versus grøn autofluorescens (lipofuscin). En port til bestemmelse af procentdelen af TIS-celler etableres baseret på ubehandlede kontrolprøver (ikke vist). Hvis der anvendes et sorteringscytometer (FACS), kan TIS-celler opsamles og placeres tilbage i kultur til yderligere in vitro-assays eller lyseres og behandles til molekylærbiologiske assays. Forkortelser: DDAO = 9H-(1,3-dichlor-9,9-dimethylacridin-2-on); DDAOG = DDAO-Galactoside; TIS = terapiinduceret ældning; FL-Ab = fluorophorekonjugeret antistof; Baf = Bafilomycin A1; SA-β-Gal = ældningsassocieret beta-galactosidase; PBS = fosfatbufferet saltvand; FACS = fluorescensaktiveret cellesortering. Klik her for at se en større version af denne figur.
| Fluorophore | Registrerer | Eks/Em (nm) | Cytometer laser (nm) | Cytometerdetektor / båndpasfilter (nm) |
| DDAOG | SA-β-Gal | 645/6601 | 640 | 670 / 30 |
| AF | Lipofuscin | < 600 | 488 | 525 / 50 |
| CV450 | Levedygtighed | 408/450 | 405 | 450 / 50 |
| PE | Antistof/overflademarkør | 565/578 | 561 | 582 / 15 |
Tabel 1: Fluoroforer og cytometer optiske specifikationer. Cytometerspecifikationer, der anvendes i denne protokol, er angivet for et instrument med i alt 4 lasere og 15 emissionsdetektorer. DDAOG detekteret ved 645/660 nm er formen af sonden spaltet af SA-β-Gal1. Uncleaved DDAOG kan udvise fluorescens på lavt niveau ved 460/610 nm, men fjernes ved vasketrin i protokollen. Forkortelser: DDAO = 9H-(1,3-dichlor-9,9-dimethylacridin-2-on); DDAOG = DDAO-Galactoside; AF = autofluorescens; PE = phycoerythrin; SA-β-Gal = ældningsassocieret beta-galactosidase.
