Senescent celler akkumuleres normalt i organismer over år under normal biologisk aldring, men kan også utvikle seg raskt i tumorceller som et svar på skade forårsaket av ulike kreftbehandlinger, inkludert stråling og kjemoterapi. Selv om det ikke lenger sprer seg, kan terapiinduserte senescent (TIS) tumorceller bidra til behandlingsresistens og drive tilbakefall 1,2,3. Faktorer utskilt av TIS-celler kan forverre tumor malignitet ved å fremme immununnvikelse eller metastase 4,5. TIS-celler utvikler komplekse, kontekstspesifikke fenotyper, endrede metabolske profiler og unike immunresponser 6,7,8. Derfor er identifisering og karakterisering av TIS-tumorceller indusert av ulike kreftbehandlingsmetoder et tema av kontinuerlig interesse for kreftforskningsmiljøet.
For å oppdage TIS-tumorceller er konvensjonelle senescensanalyser mye brukt, hovedsakelig basert på å oppdage økt aktivitet av senescensmarkørenzymet, den lysosomale beta-galaktosidase GLB19. Deteksjon ved en nesten nøytral (snarere enn sur) lysosomal pH muliggjør spesifikk påvisning av senescensassosiert beta-galaktosidase (SA-β-Gal)10. En standard SA-β-Gal-analyse som har blitt brukt i flere tiår, bruker X-Gal (5-bromo-4-chloro-3-indolyl-β-D-galactopyranoside), et blått kromogent beta-galaktosidasesubstrat, for å oppdage SA-β-Gal i faste celler ved lysmikroskopi11. X-Gal-analysen tillater kvalitativ visuell bekreftelse av TIS ved bruk av allment tilgjengelige reagenser og laboratorieutstyr. Et grunnleggende overført lysmikroskop er den eneste instrumenteringen som kreves for å evaluere tilstedeværelsen av det blå kromogenet. Imidlertid kan X-Gal-fargeprosedyren mangle følsomhet, noen ganger krever mer enn 24 timer for at farge skal utvikle seg. Farging etterfølges av lav gjennomstrømning, subjektiv scoring av individuelle senescentceller basert på å telle cellene som viser et visst intensitetsnivå av det blå kromogenet under et lysmikroskop. Siden X-Gal er celle-ugjennomtrengelig, krever denne analysen løsemiddelfikserte celler, som ikke kan gjenopprettes for nedstrømsanalyse. Ved arbeid med begrensede prøver fra dyr eller pasienter kan dette være en stor ulempe.
Forbedrede SA-β-Gal-analyser ved bruk av cellepermeante, fluorescerende enzymsubstrater, inkludert C 12-FDG (5-dodecanoylaminofluorescein Di-β-D-Galactopyranoside, grønn) og DDAOG (9H-(1,3-diklor-9,9-dimetylacridin-2-en-7-yl) β-D-Galactopyranoside, langt rød) har tidligere dukket opp i litteraturen12,13,14,15. Den kjemiske sondestrukturen og optiske egenskapene til DDAOG er vist i tilleggsfigur S1. Disse celle-permeant sonder tillater analyse av levende (snarere enn faste) celler, og fluorescerende snarere enn kromogene prober lette bruken av raske high-throughput fluorescerende analyseplattformer, inkludert høyinnhold screening instrumenter og flow cytometre. Sortering av flowcytometre muliggjør gjenoppretting av berikede populasjoner av levende senescentceller fra cellekulturer eller svulster for nedstrøms analyse (f.eks. Western blotting, ELISA eller ‘omics). Fluorescensanalyse gir også et kvantitativt signal, noe som muliggjør mer nøyaktig bestemmelse av prosentandelen av senescentceller i en gitt prøve. Ytterligere fluorescerende prober, inkludert levedyktighetssonder og fluoroformerkede antistoffer, kan lett legges til for multiplekset analyse av mål utover SA-β-Gal.
I likhet med DDAOG er C12-FDG en fluorescerende sonde for SA-β-Gal, men dens grønne fluorescerende utslipp overlapper med inneboende cellulær AF, som oppstår under aldring på grunn av akkumulering av lipofuscinaggregater i celler16. Ved å bruke den langt røde DDAOG-sonden, kan grønn cellulær AF brukes som en sekundær parameter for å bekrefte senescence17. Dette forbedrer analysepåliteligheten ved å bruke en annen markør i tillegg til SA-β-Gal, som ofte kan være upålitelig som en enkelt markør for senescence18. Siden deteksjon av endogen AF i senescentceller er en etikettfri tilnærming, er det en rask og enkel måte å utvide spesifisiteten til vår DDAOG-baserte analyse.
I denne protokollen demonstrerer vi bruken av DDAOG og AF som en rask, dual-parameter flowcytometrianalyse for identifisering av levedyktige TIS-tumorceller fra in vitro-kulturer eller isolert fra legemiddelbehandlede svulster etablert hos mus (figur 1). Protokollen bruker fluoroforer som er kompatible med et bredt spekter av standard kommersielle flowcytometrianalysatorer og sorteringsmaskiner (tabell 1). Kvantifisering av prosentandelen av levedyktige senescentceller ved bruk av standard flowcytometrianalyse er aktivert. Om ønskelig kan et valgfritt immunmerkingstrinn utføres for å evaluere celleoverflateantigener av interesse samtidig med senescens. Identifiserte senescentceller kan også berikes ved hjelp av standard fluorescensaktivert cellesortering (FACS) metodikk.
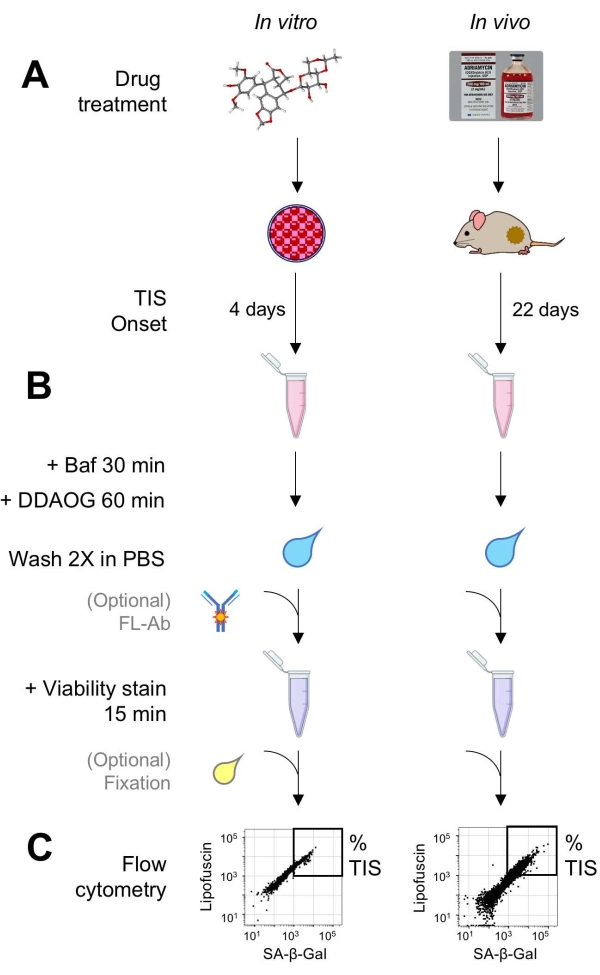
Figur 1: Eksperimentell arbeidsflyt. En skjematisk oppsummering av nøkkelpunkter i DDAOG-analysen. (A) Et TIS-induserende legemiddel tilsettes pattedyrdyrkede celler eller administreres til tumorbærende mus. Tid er da tillatt for utbruddet av TIS: for celler, 4 dager etter behandling; For mus, 22 dager totalt, med tre behandlinger hver 5. dag pluss 7 dager utvinning. Celler høstes eller svulster dissosieres til suspensjon. (B) Prøver behandles med Baf for å justere lysosomal pH for påvisning av SA-β-Gal i 30 minutter; deretter legges DDAOG-sonden til i 60 minutter for å oppdage SA-β-Gal. Prøver vaskes 2x i PBS, og en levedyktighetsflekk tilsettes kort (15 min). Eventuelt kan prøver farges med fluorescerende antistoffer i åpne fluorescenskanaler og / eller fikses for senere analyse. (C) Prøver analyseres ved hjelp av et standard flowcytometer. Levedyktige celler visualiseres i punktplott som viser rød DDAOG (indikerer SA-β-Gal) versus grønn autofluorescens (lipofuscin). En port for å bestemme prosentandelen av TIS-celler er etablert basert på ubehandlede kontrollprøver (ikke vist). Hvis et sorteringscytometer (FACS) brukes, kan TIS-celler samles og plasseres tilbake i kultur for videre in vitro-analyser eller lyses og behandles for molekylærbiologiske analyser. Forkortelser: DDAO = 9H- (1,3-diklor-9,9-dimetylacridin-2-en); DDAOG = DDAO-Galactoside; TIS = terapi-indusert senescence; FL-Ab = fluorofor-konjugert antistoff; Baf = Bafilomycin A1; SA-β-Gal = senescence-assosiert beta-galaktosidase; PBS = fosfatbufret saltvann; FACS = fluorescensaktivert cellesortering. Vennligst klikk her for å se en større versjon av denne figuren.
| Fluorophore | Oppdager | Eks/Em (nm) | Cytometer laser (nm) | Cytometer detektor / båndpassfilter (nm) |
| DDAOG | SA-β-Gal | 645/6601 | 640 | 670 / 30 |
| AF | Lipofuscin | < 600 | 488 | 525 / 50 |
| CV450 | Levedyktighet | 408/450 | 405 | 450 / 50 |
| PE | Antistoff/overflatemarkør | 565/578 | 561 | 582 / 15 |
Tabell 1: Fluoroforer og cytometer optiske spesifikasjoner. Cytometerspesifikasjoner som brukes i denne protokollen er oppført for et instrument med totalt 4 lasere og 15 utslippsdetektorer. DDAOG oppdaget ved 645/660 nm er formen av sonden spaltet av SA-β-Gal1. Uncleaved DDAOG kan vise lavt nivå fluorescens ved 460/610 nm, men fjernes ved vask trinn i protokollen. Forkortelser: DDAO = 9H- (1,3-diklor-9,9-dimetylacridin-2-en); DDAOG = DDAO-Galactoside; AF = autofluorescens; PE = phycoerythrin; SA-β-Gal = senescence-assosiert beta-galaktosidase.
