AirID-Based Proximity Labeling for Protein-Protein Interaction in Plants
Summary
Here, we present a step-by-step protocol for performing the proximity labeling (PL) experiment in cucumber (Cucumis sativus L.) using AT4G18020 (APRR2)-AirID protein as a model. The method describes the construction of a vector, the transformation of a construct through agroinfiltration, biotin infiltration, protein extraction, and purification of biotin-labeled proteins through affinity purification technique.
Abstract
In mammalian cells and plants, proximity labeling (PL) approaches using modified ascorbate peroxidase (APEX) or the Escherichia coli biotin ligase BirA (known as BioID) have proven successful in identifying protein-protein interactions (PPIs). APEX, BioID, and TurboID, a revised version of BioID have some restrictions in addition to being valuable technologies. The recently developed AirID, a novel version of BioID for proximity identification in protein-protein interactions, overcame these restrictions. Previously, AirID has been used in animal models, while the current study demonstrates the use of AirID in plants, and the results confirmed that AirID performs better in plant systems as compared to other PL enzymes such as BioID and TurboID for protein labeling that are proximal to the target proteins. Here is a step-by-step protocol for identifying protein interaction partners using AT4G18020 (APRR2) protein as a model. The methods describe the construction of vector, the transformation of construct through agroinfiltration, biotin transformation, extraction of proteins, and enrichment of biotin-labeled proteins through affinity purification technique. The results conclude that AirID is a novel and ideal enzyme for analyzing PPIs in plants. The method can be applied to study other proteins in plants.
Introduction
Various cellular proteins work under the biologically regulatory system, and protein-protein interactions (PPIs) are a part of this system and the basis of many cellular processes. Besides PPIs, the function of natural proteins is post-translationally promoted via various modifications such as the formation of complex, ubiquitination, and phosphorylation. Therefore, studying PPIs is significant to understanding the possible function of target proteins. PPIs have been carried out using various technologies such as mass spectrometry analysis after immunoprecipitation (IP-MS analysis)1, yeast two-hybrid system (Y2H)2, also cell-free based arrays3. These methods explored various vital findings in the field of research. However, these methods have some drawbacks; for example, Y2H is a time-consuming, expensive strategy that necessitates building the target species' Y2H library.
Additionally, the Y2H technique uses yeast, a heterologous single-cell eukaryotic organism, which could not accurately reflect the cellular state of higher eukaryotic cells. The IP-MS is unsuitable for high hydrophobicity proteins and shows low efficiency in capturing weak PPIs. Various essential proteins in plants such as nucleotide-binding domain and leucine-rich repeat-containing (NLR) proteins and receptor-like kinases (RLKs) are expressed at a low level and mostly interact with other proteins transiently; therefore, using these methods is insufficient for understanding the mechanisms underlying the regulation of these proteins3.
A new technique called proximity biotinylation (PB) helps researchers identify PPIs. PB depends on PL enzymes, which attach to the protein of interest (POI), and when partner protein comes near POI, the PL attaches a chemical biotin tag to the partner protein. Further, the tagged protein can be identified and can quickly know which partner protein attaches to the target protein5. Previous studies proved that BioID and TurboID are successful tools for PPIs, especially in plants, but they have certain limitations4. BioID needs a high level of biotin for labeling partner proteins, which takes more than 16 h. Compared to BioID, the TurboID is more beneficial as it labels protein in 10 min and can label the partner protein at room temperature (RT). It is also toxic to cells in certain conditions and tags those proteins that do not show interaction with the protein of interest.
To overcome these issues, AirID, developed by Kido et al., is more efficient than the rest of the labeling enzymes, although the sequence similarity is 82% between BioID and AirID5. To check the efficiency of AirID, we conducted an experiment by using a POI with known associates. This experiment confirmed that AirID could undoubtedly label associated proteins in plant cells. AirID is a valuable enzyme for analyzing PPIs in vitro and in cells. It creates less toxicity and is less erroneous in time taken processes than TurboID to tag non-partners, leading to killing the cell. It demonstrates that AirID is more competitive than other labeling enzymes for proximity biotinylation. It is more accurate, has more potential in time-taking processes, and less toxic in vitro and in living cells. The current protocol describes the identification of interacting proteins of APRR2 using AirID as a PL enzyme; furthermore, the method can be applied to other proteins to investigate PPIs in plant species.
Protocol
1. Preparation of plant material
- Cucumis sativus (Cucumber) is employed for experimental analysis. Put the seeds in water, incubate at 50 °C for 20 min, and then place the seeds on filter paper on a Petri plate for 12-16 h.
- Afterward, transfer the seeds to pots containing soil (purchased commercially) and grow in a climatic chamber at 23 °C temperature and 16 h light and 8 h dark photoperiod.
- Maintain the plants in the climate chamber for 3-4 weeks until the plants reach 3 or 4 leaf stages for successful agroinfiltration.
2. Making AirID construct
- Use APRR2 as a target protein and construct APRR2-AirID under the Cauliflower mosaic virus 35S promoter (p35S: APRR2-AirID). Introduce it into the Gateway-compatible binary vector pEarleyGate1006 (provided by Dr. Kathrin Schrick, Kansas State University), and synthesize the PL enzyme directly according to the sequence provided in Supplementary File 1.
3. Preparation of competent cells
- Preparation of DH5α competent cells
- Inoculate the 2 mL of LB (10 g/L of tryptone, 5 g/L of yeast extract, and 10 g/L of NaCl; pH = 7.0.) with E.Coli DH5α cells (directly from the frozen stock without thawing) and grow overnight (O/N) at 37 °C.
- Freshly inoculate 0.1%-0.5% inoculum from the O/N culture into 100 mL of LB Medium and grow the cells until OD600 reaches between 0.4-0.6, and then Incubate the culture at 4 °C for 30 min.
- Take the culture into two 50 mL centrifuge tubes and spin at 2500 x g for 5 min at 4 °C.
- Discard the supernatant, resuspend the cells in 10 mL of 0.1 M ice cold CaCl2, and set on ice for 15 min. Pellet the cells with a 2500 x g spin for 5 min at 4 °C.
- Resuspend the cells in 1 mL of ice-cold 0.1 M CaCl2 + 20% glycerol, aliquot 100 µL into each 1.5 mL tube, and store them immediately at -80 °C.
- Preparation of GV3101 competent cells
- Inoculate 2 mL of LB (containing 50 µg/mL gentamycin and 25 µg/mL rifampicin) with a single GV3101 colony. Incubate the culture O/N at 28 °C while shaking at 250 rpm.
- Inoculate 200 mL of LB with 1 mL of a saturated overnight culture. Shake the culture at 250 rpm while incubating it at 28 °C until the OD600 is equal to 0.5.
- Transfer the culture to four pre-chilled sterile 50 mL centrifuge tubes and set on ice for 30 min.
- Pellet the cells at 2500 x g for 10 min at 4 °C. Discard the supernatant and place the pellets on ice.
- Resuspend the cells in 10 mL of 0.1 M ice cold CaCl2 solution. Pool the cells together into one pre-chilled 50 mL Oakridge tube.
- Pellet the cells at 1,000 x g for 5 min at 4 °C. Discard the supernatant and resuspend the cells in 10 mL of ice cold CaCl2 solution. Set on ice for 30 min.
- Pellet the cells at 1,000 x g for 5 min at 4 °C. Discard the supernatant and resuspend the cells in 2 mL of 0.1 M ice cold CaCl2 solution.
- Dispense the cells into a 50 µL aliquot in pre-chilled sterile polypropylene tubes. Store the cells at -80 °C.
4. Agroinfiltration
- First, transfer the plasmid to agrobacterium
- Transfer 2.5 µL of the plasmid generated from step 2.1 to the competent cells of the agrobacterium tumefaciens GV3101 strain and incubate on ice for 30 min.
- Transfer the tube containing the plasmid and the competent cells to liquid nitrogen for 3 min.
- Heat shock at 37 °C for 5 min in an incubator, and then put the tube on ice for 2 min.
- Add 1,000 µLof LB medium (10 g/L of tryptone, 5 g/L of yeast extract, and 10 g/L of NaCl; pH = 7.0) to the agrobacteriumand incubate at 30 °C and 118 rpm for 1 h.
- Centrifuge at 3,000 x g for 2 min at 4 °C.
- Discard the upper 800 µL of the solution and mix the remaining solution. Then, plate it on LB (as mentioned in step 4.1.4 added with agar and 50 µg/mL Kanamycin for plasmid and 50 µg/mL gentamycin and 25 µg/mL rifampicin for GV3010 competent cells). Incubate the plates at 30 °C for 48 h.
NOTE: It is better to pick some colonies and perform PCR to confirm the gene of interest7.
- Pick up some bacterial colonies from the plates and put them in LB media plus appropriate antibiotics (see step 4.1.6). Incubate at 30 °C and 218 rpm for 36-48 h.
- Centrifuge the cells at 3,000 x g for 2 minat 4 °C. Resuspend the cells to OD600 = 1.0 in Agroinfiltration buffer (10 mM MgCl2, 10 mM MES, pH = 5.6, 250 µM acetosyringone).
- Infiltrate the inoculum (generated from step 4.3) in the (abaxial) epidermis using a 1 mL needleless syringe.
NOTE: It is better to infiltrate the entire leaf and choose to replicate. For 4-5 plants, use one construct. For each leaf, 1.5 mL of resuspended agrobacteria is sufficient. - Maintain the plants for 36 h in the climate chamber with a 16 h light (about 75 µmol·m-2·s-1)and 8 h dark photoperiod at 23 °C.
- After 36 h of construct infiltration, infiltrate 1 mL of 5 µM Biotin (in 10 mM MgCl2 solution) into the already infiltered leaves of the construct.
- Maintain the treated plants for an additional 4-12 h before harvesting the leaf tissue.
NOTE: According to the previous study, the target protein peaks at 36 h, so choose 36 hpi biotin infiltrations4,6. The incubation duration for biotin depends on the target study, but according to the current experiment, an 8 h incubation period is more suitable for labeling proteins.
5. Collection of samples
NOTE: All materials for sample collection should be sterile to avoid keratin contamination, and all the protocol steps should be performed in a contamination-free environment.
- Cut the infiltrated leaves and quickly transfer them to liquid nitrogen to avoid protein degradation.
- Perform the pre-detection of protein through immunoblot analyses6; this is highly recommended before Co-immunoprecipitation (Co-IP).
6. Total protein extraction from leaf
- Grind the leaves using a pestle and mortar, quickly add 2 mL of 1x PBS-BSA PH 7.4 and grind slowly.
- Take a 15 mL conical tube, place a quick filtration material filter on top of the conical tube, transfer the sample mix to the tube through the quick filtration material filter, and keep it on ice.
- Transfer the samples to a 2 mL tube and add 1% protein inhibitor cocktail.
- Mix the contents by turning upward and downward 7-8 times and centrifuge at 1,000 x g for 2 min at 4 °C.
- Transfer the upper solvent of samples to new 2 mL tubes, add 10% β-D-maltoside (β-D.M), and place on ice for 5 min. Centrifuge at 20,000 x g for 10 min at 4 °C.
7. Equilibrate the desalting column
- Centrifuge the column at 1,000 x g for 1 min at 4 °C.
NOTE: Mark one side of the column and ensure that the marked side faces outward during all centrifugation processes. - Remove the top and bottom cover of the column and centrifuge at 1,000 x g for 2 min at 4 °C.
- Discard the liquid solution from the collection tube of the column and add 5 mL of 1x PBS buffer for washing.
- Centrifuge the column at 1,000 x g for 2 min at 4 °C.
- Repeat the steps 7.3 and 7.4 at least five times.
8. Magnetic beads washing
- Take a 1.5 mL tube and add 50 µL of streptavidin-C1-conjugated magnetic beads.
- Add 1 mL of 1x PBS-BSA for washing, mix thoroughly and place on the magnet stand for 3 min. Discard the supernatant.
- Repeat step 8.2 at least three times.
- After each washing, place the tube on the magnetic rack to adsorb the beads toward one side of the tube to remove the washing buffer.
9. Enrichment of biotinylated proteins
- Add 2 mL of the samples to the column and centrifuge at 1,000 x g for 8 min at 4 °C.
- Add 1 mL of the desalted protein extract to 50 µL of streptavidin-C1-conjugated magnetic beads to equilibrate them.
- Place the tube on the rotator at normal speed so the solution mixes thoroughly and incubates at room temperature (RT) for 30 min.
- Place the beads on the magnetic rack for 3 min at RT, or until they gather on one side of the tube, to gently remove the supernatant.
- Add 1 mL of wash buffer I (2% SDS in water), rotate at RT for 2 min on a rotator and repeat step 9.4.
- Place the tube on the rotator at RT for 2 min after adding 1 mL of wash buffer II (50 mM HEPES: pH = 7.5, 500 mM NaCl, 1 mM EDTA, 0.1% deoxycholic acid [w/v], and 1% Triton X-100). Repeat step 9.4.
- Add 1 mL of wash buffer III (10 mM Tris-HCl: pH = 7.4, 250 mM LiCl, 1 mM EDTA, 0.1% deoxycholic acid [w/v], 1% NP40 [v/v]), and rotate using a shaker for 2 min at RT. Then, repeat step 9.4.
- To remove the detergent, add 1.7 mL of 50 mM Tris-HCl (pH = 7.5) and repeat step 9.4. Repeat this step one more time.
- Wash the beads six times for 2 min each in 1 mL of 50 mM ammonium bicarbonate buffer at RT and repeat step 9.4.
- Add 50 µL of the protein extract containing 50 mM Tris-HCl (pH 8.0), 12% (w/v) sucrose (Suc), 2% (w/v) lithium lauryl sulfate, and 1.5% (w/v) dithiothreitol to the beads, heat shock in the incubator at 100 °C for 5 min and follow step 9.4. Store the supernatant at -80 °C for LC-MS/MS analysis.
Representative Results
According to previous research, the cucumber gene APRR2 is the candidate gene that controls white immature fruit color8. Here, a protocol was developed using AirID as a proximity labeling enzyme to find the interacting partner protein of APRR2 in cucumber. The construct was transferred to the cucumber leaves, and after 36 h post infiltration, biotin was transferred. After 48 h the samples were taken for western blot analysis to confirm the successful transformation. The proteins were extracted as mentioned above in the protocol for western blot. The representative data in Figure 1 indicated the protein expression and biotinylation in the infiltrated cucumber leaves, which were the expected findings based on the newly modified technique. The results showed a higher expression level of labeled proteins and extra bands as compared to the control. This confirms that AirID successfully tagged the target proteins of the gene of interest (GOI). Further, the results were also confirmed using anti-flag and anti-mouse antibodies, which successfully showed the target band with anti-flag antibody (Figure 2). After confirmation through western blot analysis, we further performed Co-IP using the protocol mentioned above, and the infiltrating leaves' biotinylated proteins were successfully enriched for mass spectrometry analysis, as shown in Figure 3. After enriching biotinylated proteins with streptavidin C1 conjugated magnetic beads, multiple proteins of different sizes were observed, and western blot analysis revealed smeared bands (Figure 3).
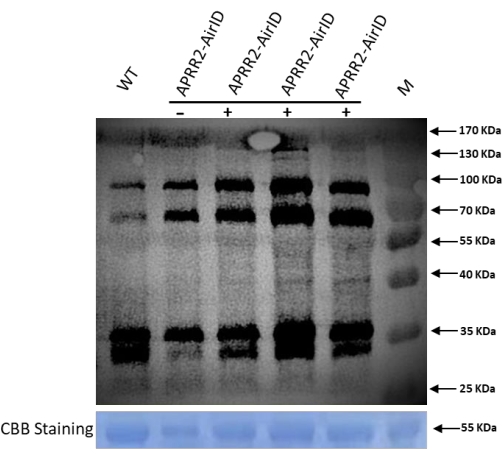
Figure 1: Western blot analysis of proteins after transformation to leaves to confirm the function of AirID using streptavidin-HRP. The figure confirms that biotinylated proteins have four independent replicates for the construct transformed samples. Wild-type leaves were used as a control. For each replicate, 10 µL of the samples were loaded for western blot analysis. Streptavidin-HRP (1:6000 dilution) was used to analyze biotinylated proteins in different samples. Please click here to view a larger version of this figure.
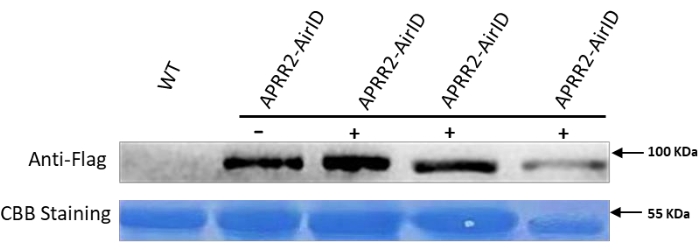
Figure 2: Western blot analysis of proteins after transformation to leaves to detect the Taq sequence. The 3x-Flag tag sequence was fused with the gene of interest to confirm the successful transformation. The taq sequence was identified through western blot analysis in all the transformed samples. Wild-type cucumber leaf samples were used as a control. A total 10 µL of the samples were loaded for western blot analysis, and the results were confirmed using anti-flag as a first antibody at dilution of 1:3000 and anti-mouse as a second antibody at dilution 1:10000. Please click here to view a larger version of this figure.
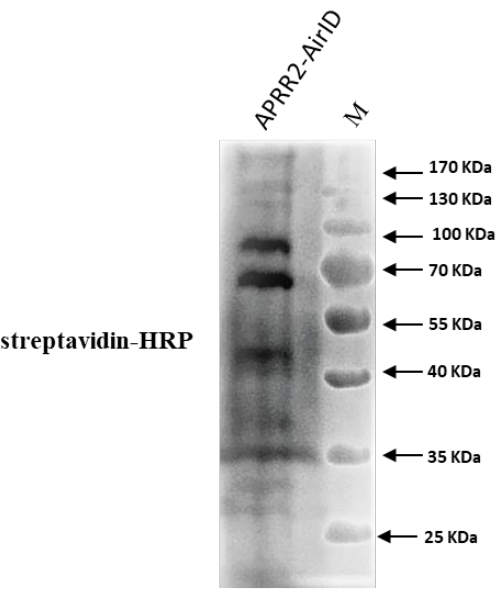
Figure 3: Immunoblot analysis of protein extracts. After Co-immunoprecipitation (Co-IP), Western blot analysis of biotinylated proteins in the agroinfiltrated leaves of Cucumis sativus (Cucumber) with streptavidin-HRP was performed. Please click here to view a larger version of this figure.
Supplementary File 1. Please click here to download this File.
Discussion
In the current experiment, AirID was used for proximity labeling, which Kido et al. developed through an algorithm of ancestral enzyme reconstruction using a large genome dataset and five conventional BirA enzymes5. Random mutations were used in traditional evolutionary protein engineering to enhance activity9,10 as random mutations cannot produce dynamic sequence changes. Compared to other PL enzymes, AirID has several advantages. Previously this PL method was only applicable to animal systems. In this study, it was used to investigate protein-protein interactions in plants. The protocol outlines how to set up the AirID-based PL in plants step by step, including preparing leaf samples, removing free biotin, quantifying extracted proteins, and enriching biotinylated proteins.
Using the well-established Agrobacterium-mediated transient expression in the Cucumber, this approach is used to find PPIs of the target protein of interest in Cucumber. The transient expression may result in overexpression of the fusion proteins. The AirID fusion must also be tested to ensure that it does not interfere with the function of the target protein of interest, which is another crucial consideration. However, while PL provides several advantages over standard IP-MS techniques for detecting transitory or weak PPIs, it also has some limitations. First and foremost, discovering a candidate interaction protein does not automatically imply a direct or indirect interaction with the bait protein but instead reflects near proximity to the bait protein9. PPIs can be further verified in vivo using independent assays (e.g., co-immunoprecipitation, bimolecular fluorescence complementation (BiFC), or in vitro GST-pull down assay, which can be carried out to verify PPIs further.
The study found that the proximal biotinylation activity of AirID was significantly lower than that of TurboID. In vitro and in cells, TurboID had the highest proximal biotinylation activity. This enzyme may be utilized within 10 min to biotinylate9,11. However, in cells treated for a prolonged incubation of over 6 h and higher biotin concentrations, highest TurboID activity led to extra biotinylation on unexpected proteins, such as tubulin and GFP (such as 50 mM biotin). In contrast to being used as a proximal biotinylation enzyme for PPIs, TurboID was first reported as a biotin-labeling enzyme4,6,5,7. If TurboID were to evaluate PPIs, it would function best under limiting conditions, such as time duration is short, about 1 h in cells. The AirID, biotinylation of tubulin and GFP were not found under the same conditions as those observed for TurboID biotinylation. AirID-fusion proteins were found to be capable of biotinylation of each well-known interactor in both transients, as demonstrated by the streptavidin-pull down assay and the LC-MS/MS analysis5. In low ATP concentrations (1 mM), the formation of biotinoyl-5-AMP was stronger for AirID than for TurboID. It favors lower concentrations of biotin (with 5 mM biotin or without biotin supplement) than TurboID, which prefers higher concentrations. In addition, an examination of biotinylation sites using LC-MS/MS revealed that AirID biotinylation occurred with no unique sequence preference on a nearby Lys residue, indicating that the biotinylation process was non-preferentia5. Although the sequence similarity between BioID and AirID is 82%, AirID showed high biotinylation activity against interacting proteins. Kido et al., 2020 found that AirID can analyze protein-protein interactions in vitro and in cells. Their findings suggest that biotinylation dependent on AirID may be useful for PPI analysis of chemical compounds. The identification of in vivo partners of target proteins is crucial for understanding biological functions12,13, and it has revealed new PPIs, so in vivo proximity biotinylation using BioID has been used in numerous studies. Even when biotin was supplemented, stable expression of AirID did not result in cell-growth inhibition, indicating that the expression of AirID-fusion proteins would be very low-toxic5. AirID is projected to improve PPI-dependent biotinylation accuracy in combination; in summary, it is concluded that AirID is ideal for PPI analysis in plants.
Disclosures
The authors have nothing to disclose.
Acknowledgements
This work was supported by the National Natural Science Foundation of China (Grant No. 32000197 to X.H.), the Special Financial Grant from the China Postdoctoral Science Foundation (Grant No. 2019T120467 to X.H.)
Materials
| Acetosyringone | Beijing solaribo science and technology Co.Ltd | S1519 | |
| Acryl/Bis 30% solution | Sangon Biotech (Shanghai) Co.Ltd | 1510KA4528 | |
| Agar | BioFroxx GmbH | D64683 | |
| Agarose | tsingke (Shanghai) Co.Ltd | TSJ001 | |
| Ammonium bicarbonate | Sangon Biotech (Shanghai) Co.Ltd | G313BA0018 | |
| Biotin | BBI life Sciences | G908BA0012 | |
| CaCl2 | BBI life Sciences | E209BA0008 | |
| Competent cells GV3101 | Made in the current experiment | ||
| Desalting column | Thermo scientific | WC321753 | |
| Deoxycholic acid | Sangon Biotech (Shanghai) Co.Ltd | G818BA0029 | |
| DH5α competent cells | Made in the current experiment | E.coli DH5α | |
| β-D-maltoside | Beijing Scolario Science and Tech Co.Ltd | S818 | |
| EDTA | Sangon Biotech (Shanghai) Co.Ltd | E104BA0029 | |
| Glycine | Sangon Biotech (Shanghai) Co.Ltd | 161BA0031 | |
| HEPES | Beijing solaribo science and technology Co.Ltd | H8090 | |
| LiCl | Sangon Biotech (Shanghai) Co.Ltd | H209BA0003 | |
| MES | Beijing solaribo science and technology Co.Ltd | M8019 | |
| MiraCloth | EMD Milipore Corp/MERCK kgAa Darmstadt, Germenay | 3429963 | Quick filtration material filter |
| MgCl2 | Beijing solaribo science and technology Co.Ltd | 20200819 | |
| NaCl | Sangon Biotech (Shanghai) Co.Ltd | H324BA0003 | |
| NP40 | Sangon Biotech (Shanghai) Co.Ltd | N8030 | |
| Protein inhibitor cocktail | Beijing Scolario Science and Tech Co.Ltd | S3450 | |
| PVDF | BIO-RAD | 5820172 | |
| SDS | Beijing Scolario Science and Tech Co.Ltd | S1015 | |
| Silwet | Sangon Biotech (Shanghai) Co.Ltd | S9430 | |
| Streptavidin-C1-conjugated magnetic beads | Enriching Biotechnology | 7E511E1 | Magnetic beads |
| TEMED | Servicebio | G2056 | |
| Triton X-100 | Sangon Biotech (Shanghai) Co.Ltd | GB03BA007 | |
| Tris-HCl | Sangon Biotech (Shanghai) Co.Ltd | F828BA0020 | |
| Tryptone | Thermo scientific | LP0042 |
References
- Han, J., et al. The identification of novel protein-protein interactions in liver that affect glucagon receptor activity. PloS One. 10 (6), 0129226 (2015).
- Zhao, S., et al. Screening and identification of host proteins interacting with Theileria annulata cysteine proteinase (TaCP) by yeast-two-hybrid system. Parasites & Vectors. 10 (1), 536 (2017).
- Zhang, Y., et al. A highly efficient agrobacterium-mediated method for transient gene expression and functional studies in multiple plant species. Plant Communications. 1 (5), 100028 (2020).
- Zhang, Y., et al. TurboID-based proximity labeling for in planta identification of protein-protein interaction networks. Journal of Visualized Experiments. (159), e60728 (2020).
- Kido, K., et al. AirID, a novel proximity biotinylation enzyme, for analysis of protein-protein interactions. eLife. 9, 54983 (2020).
- Hu, X., et al. Overexpressing 7-hydroxymethyl chlorophyll a reductase alleviates non-programmed cell death during dark-induced senescence in intact Arabidopsis plants. Biomolecules. 11 (8), 1143 (2021).
- Ammiraju, J., et al. The Oryza bacterial artificial chromosome library resource: construction and analysis of 12 deep-coverage large-insert BAC libraries that represent the 10 genome types of the genus Oryza. Genome Research. 16 (1), 140-147 (2006).
- Liu, C., et al. Albino Leaf 2 is involved in the splicing of chloroplast group I and II introns in rice. Journal of Experimental Botany. 67 (18), 5339-5347 (2016).
- Branon, T. C., et al. Efficient proximity labeling in living cells and organisms with TurboID. Nature Biotechnology. 36 (9), 880-887 (2018).
- Nakano, S., Niwa, M., Asano, Y., Ito, S. Following the evolutionary track of a highly specific l-Arginine oxidase by reconstruction and biochemical analysis of ancestral and native enzymes. Applied and Environmental Microbiology. 85 (12), 00459 (2019).
- Gingras, A. -. C., Abe, K. T., Raught, B. Getting to know the neighborhood: using proximity-dependent biotinylation to characterize protein complexes and map organelles. Current Opinion in Chemical Biology. 48, 44-54 (2019).
- Odeh, H. M., Coyaud, E., Raught, B., Matunis, M. J. The SUMO-specific isopeptidase SENP2 is targeted to intracellular membranes via a predicted N-terminal amphipathic α-helix. Molecular Biology of the Cell. 29 (15), 1878-1890 (2018).
- Motani, K., Kosako, H. Activation of stimulator of interferon genes (STING) induces ADAM17-mediated shedding of the immune semaphorin SEMA4D. Journal of Biological Chemistry. 293 (20), 7717-7726 (2018).
