En 2011, los investigadores propusieron un método computacional y desarrollaron una pieza correspondiente de software para el diseño automático de circuitos genéticos sintéticos digitales1. Un usuario tenía que especificar el número de entradas (tres o cuatro) y rellenar la tabla de verdad del circuito; Esto proporcionó toda la información necesaria para derivar la estructura del circuito utilizando técnicas de la electrónica. La tabla de verdad se tradujo a dos fórmulas booleanas a través del método 2 del mapa de Karnaugh. Cada fórmula booleana está hecha de cláusulas que describen operaciones lógicas (suma o multiplicación) entre (parte de) las entradas del circuito y sus negaciones (los literales). Las cláusulas, a su vez, se suman (OR) o se multiplican (AND) para calcular la salida del circuito. Cada circuito se puede realizar de acuerdo con cualquiera de sus dos fórmulas correspondientes: una escrita en forma POS (producto de sumas) y la otra en representación SOP (suma de productos). El primero consiste en una multiplicación de cláusulas (es decir, puertas booleanas) que contienen una suma lógica de los literales. Este último, en cambio, es una suma de cláusulas donde los literales se multiplican.
Los circuitos eléctricos se pueden realizar, en una placa de pruebas, conectando físicamente diferentes puertas juntas. La corriente eléctrica permite el intercambio de señales entre puertas, lo que conduce al cálculo de la salida.
En biología, la situación es más compleja. Una puerta booleana se puede realizar como una unidad de transcripción (TU; es decir, la secuencia “promotor-codificante de la región-terminador” dentro de las células eucariotas), donde la transcripción o la traducción (o ambas) están reguladas. Así, al menos dos tipos de moléculas establecen un cableado biológico: las proteínas del factor de transcripción y los ARN antisentido no codificantes1.
Un circuito digital de genes está organizado en dos o tres capas de puertas, a saber: 1) la capa de entrada, que está hecha de puertas SÍ (tampón) y NO y convierte los productos químicos de entrada en moléculas de cableado; 2) la capa interna, que consiste en tantas TU como cláusulas hay en la fórmula booleana correspondiente. Si el circuito está diseñado de acuerdo con la fórmula SOP, cada cláusula en la capa interna producirá la salida del circuito (por ejemplo, fluorescencia) en una llamada arquitectura de salida distribuida. Si se utiliza la fórmula del producto de suma (POS), entonces se requiere una capa final 3), que contendrá una sola puerta multiplicativa que recoge las moléculas de cableado de la capa interna.
En general, en biología sintética, se pueden diseñar muchos esquemas diferentes para el mismo circuito. Difieren en el número y el tipo de TU y moléculas de cableado. Para elegir la solución más fácil de implementar en celdas de levadura, cada diseño de circuito se asocia con una puntuación de complejidad S, definida como
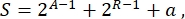
donde A representa el número de activadores, R representa el número de represores y a es la cantidad de moléculas de ARN antisentido. Si los activadores o represores están ausentes del circuito, su contribución a S es cero. Por lo tanto, es más difícil realizar un esquema de circuito en el laboratorio (S alta) cuando requiere un alto número de factores de transcripción ortogonales. Esto significa que los nuevos activadores y represores deben diseñarse de novo para realizar el cableado completo dentro de los circuitos digitales. En principio, las nuevas proteínas de unión al ADN se pueden ensamblar utilizando las proteínas Zinc Finger3 y los efectores TAL4 como plantillas. Sin embargo, esta opción parece demasiado ardua y lenta; por lo tanto, uno debe confiar principalmente en pequeños ARN y regulación de traducción para finalizar circuitos genéticos complejos.
Originalmente, este método fue desarrollado para fabricar circuitos digitales en bacterias. De hecho, en las células eucariotas, en lugar de ARN antisentido, es más adecuado hablar de microARN (miARN) o pequeños ARN interferentes (siRNAs)5. Sin embargo, la vía del ARNi no está presente en la levadura S. cerevisiae. Por lo tanto, uno debe optar por redes completamente transcripcionales. Supongamos que un circuito necesita cinco activadores y cinco represores; su puntuación de complejidad sería S = 32. La complejidad del circuito puede reducirse reemplazando los 10 factores de transcripción con un solo dCas96 (Cas9 deficiente en nucleasa) fusionado a un dominio de activación (AD). Como se muestra en7, dCas9-AD funciona como un represor en la levadura cuando se une a un promotor entre la caja TATA y el TSS (sitio de inicio de la transcripción) y como un activador cuando se une bien aguas arriba de la caja TATA. Por lo tanto, se pueden reemplazar 10 factores de transcripción con una sola proteína de fusión dCas9-AD y 10 sgRNAs (ARN guía únicos) para una puntuación de complejidad total de S = 11. Es rápido y fácil sintetizar diez sgRNAs, mientras que, como se comentó anteriormente, el ensamblaje de 10 proteínas exigiría un trabajo mucho más largo y complicado.
Alternativamente, se podrían usar dos proteínas dCas ortogonales (por ejemplo, dCas9 y dCas12a): una para fusionarse con un AD y la otra desnuda o en combinación con un dominio de represión. La puntuación de complejidad aumentaría en una sola unidad (S = 12). Por lo tanto, los sistemas CRISPR-dCas son la clave para la construcción de circuitos digitales de genes muy intrincados en S. cerevisiae.
Este documento caracteriza profundamente la eficiencia de los represores y activadores basados en dCas9 y dCas12a en levadura. Los resultados muestran que no demandan una gran cantidad de sgRNA para optimizar su actividad, por lo que los plásmidos episomales se evitan preferentemente. Además, los activadores basados en dCas9 son mucho más efectivos cuando se usa un ARN de andamio (scRNA) que recluta copias del VP64 AD. Por el contrario, dCas12a funciona bien cuando se fusiona directamente con el fuerte VPR AD. Además, un promotor activado sintético exige un número variable de sitios diana, dependiendo de la configuración del activador (por ejemplo, tres cuando se utiliza dCas12a-VPR, seis para dCas9-VP64 y solo uno con dCas9 y un scRNA). Como represor, dCas12a parece más incisivo cuando se une a la región codificante que al promotor.
Sin embargo, como inconveniente, CRISPR-dCas9 / dCas12a no interactúan directamente con los productos químicos. Por lo tanto, es posible que no sean útiles en la capa de entrada. Por esta razón, se han investigado diseños alternativos de puertas booleanas que contienen proteínas anti-CRISPR (Acrs). Acrs actúan sobre las proteínas (d)Cas e inhiben su funcionamiento8. Por lo tanto, son un medio para modular la actividad de los sistemas CRISPR-(d)Cas. Este artículo analiza a fondo las interacciones entre Acrs tipo II y (d)Cas9, así como Acrs tipo V y (d)Cas12a en S. cerevisiae. Dado que los Acr son mucho más pequeños que las proteínas Cas, se construyó una puerta NOT que responde al estrógeno β-estradiol fusionando el dominio de unión hormonal del receptor de estrógeno humano9-HBD (hER) con AcrIIA4. Además, se realizaron un puñado de puertas SÍ y NO que expresaban dCas12a (-AD) constitutivamente y AcrVA en la inducción con galactosa. En la actualidad, estas puertas sirven solo como prueba de concepto. Sin embargo, también representan el primer paso hacia un replanteamiento profundo del algoritmo para llevar a cabo el diseño automático computacional de circuitos digitales de genes sintéticos en células de levadura.
