2011 yılında, araştırmacılar hesaplamalı bir yöntem önerdiler ve dijital sentetik gen devrelerinin otomatik tasarımı için karşılık gelen bir yazılım parçası geliştirdiler1. Bir kullanıcının giriş sayısını (üç veya dört) belirtmesi ve devre doğruluk tablosunu doldurması gerekiyordu; Bu, elektronikten teknikler kullanarak devre yapısını türetmek için gerekli tüm bilgileri sağladı. Doğruluk tablosu, Karnaugh harita yöntemi2 aracılığıyla iki Boole formülüne çevrildi. Her Boole formülü, devre girişleri ve bunların olumsuzlamaları (değişmez değerler) arasındaki (bir kısmı) mantık işlemlerini (toplama veya çarpma) tanımlayan yan tümcelerden oluşur. Yan tümceler, sırasıyla, devre çıkışını hesaplamak için ya toplanır (OR) ya da çarpılır (AND). Her devre, karşılık gelen iki formülden herhangi birine göre gerçekleştirilebilir: biri POS (toplamların ürünü) formunda, diğeri SÇP (ürünlerin toplamı) temsilinde yazılmıştır. İlki, değişmez değerlerin mantıksal bir toplamını içeren cümleciklerin (yani, Boole kapıları) çarpımından oluşur. İkincisi, aksine, değişmez değerlerin çarpıldığı bir cümle toplamıdır.
Elektrik devreleri, bir breadboard üzerinde, farklı kapıları fiziksel olarak birbirine bağlayarak gerçekleştirilebilir. Elektrik akımı, kapılar arasında sinyal alışverişine izin verir ve bu da çıkışın hesaplanmasına yol açar.
Biyolojide durum daha karmaşıktır. Bir Boole kapısı, transkripsiyon veya translasyonun (veya her ikisinin) düzenlendiği bir transkripsiyon birimi (TU; yani, ökaryotik hücreler içindeki “promotör-kodlayıcı bölge-sonlandırıcı” dizisi) olarak gerçekleştirilebilir. Böylece, en az iki tür molekül biyolojik bir kablolama kurar: transkripsiyon faktörü proteinleri ve kodlamayan, antisens RNA’lar1.
Bir gen dijital devresi, iki veya üç kapı katmanı halinde düzenlenir, yani: 1) EVET (tampon) ve NOT kapılarından oluşan ve giriş kimyasallarını kablolama moleküllerine dönüştüren giriş katmanı; 2) karşılık gelen Boole formülünde yan tümceler olduğu kadar TU’dan oluşan iç katman. Devre SÇP formülüne göre tasarlanmışsa, iç katmandaki her cümle, dağıtılmış çıkış mimarisi olarak adlandırılan bir devrede devre çıkışını (örneğin, floresan) üretecektir. Toplam (POS) formülünün ürünü kullanılırsa, kablolama moleküllerini iç katmandan toplayan tek bir çarpma kapısı içerecek olan 3) son katman gereklidir.
Genel olarak, sentetik biyolojide, aynı devre için birçok farklı şema tasarlanabilir. Hem TU’ların hem de kablolama moleküllerinin sayısı ve türü bakımından farklılık gösterirler. Maya hücrelerinde uygulanacak en kolay çözümü seçmek için, her devre tasarımı, şu şekilde tanımlanan bir karmaşıklık skoru S ile ilişkilendirilir:
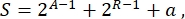
burada A, aktivatörlerin sayısını, R, baskılayıcıların sayısını ve A, antisens RNA moleküllerinin miktarını temsil eder. Aktivatörler veya baskılayıcılar devrede yoksa, S’ye katkıları sıfırdır. Bu nedenle, çok sayıda ortogonal transkripsiyon faktörü gerektirdiğinde laboratuarda bir devre şeması (yüksek S) gerçekleştirmek daha zordur. Bu, dijital devrelerin içindeki tüm kablolamayı gerçekleştirmek için yeni aktivatörlerin ve baskılayıcıların de novo olarak tasarlanacağı anlamına gelir. Prensip olarak, yeni DNA bağlayıcı proteinler, şablon olarak Çinko Parmak proteinleri3 ve TAL efektörleri4 kullanılarak birleştirilebilir. Ancak, bu seçenek çok zorlu ve zaman alıcı görünüyor; bu nedenle, karmaşık gen devrelerini sonlandırmak için çoğunlukla küçük RNA’lara ve çeviri düzenlemesine güvenilmelidir.
Başlangıçta, bu yöntem bakterilerde dijital devreler üretmek için geliştirilmiştir. Gerçekten de, ökaryotik hücrelerde, antisens RNA’lar yerine, mikroRNA’lardan (miRNA’lar) veya küçük müdahale eden RNA’lardan (siRNA’lar)5 bahsetmek daha uygundur. Bununla birlikte, RNAi yolu maya S. cerevisiae’de mevcut değildir. Bu nedenle, kişi tamamen transkripsiyonel ağları tercih etmelidir. Bir devrenin beş aktivatöre ve beş baskıcıya ihtiyacı olduğunu varsayalım; karmaşıklık puanı S = 32 olacaktır. Devre karmaşıklığı, 10 transkripsiyon faktörünün bir aktivasyon alanına (AD) kaynaşmış tek bir dCas96 (nükleaz eksikliği Cas9) ile değiştirilmesiyle azaltılabilir. 7’de gösterildiği gibi, dCas9-AD, TATA kutusu ile TSS (transkripsiyon başlangıç bölgesi) arasında bir promotör bağlarken mayada bir baskılayıcı olarak ve TATA kutusunun yukarı akışını iyi bağlarken bir aktivatör olarak çalışır. Böylece, 10 transkripsiyon faktörünü, tek bir dCas9-AD füzyon proteini ve 10 sgRNA (tek kılavuz RNA’lar) ile S = 11’lik toplam karmaşıklık skoru için değiştirebiliriz. On sgRNA’yı sentezlemek hızlı ve kolaydır, oysa daha önce yorumlandığı gibi, 10 proteinin montajı çok daha uzun ve daha karmaşık bir çalışma gerektirecektir.
Alternatif olarak, biri iki ortogonal dCas proteini (örneğin, dCas9 ve dCas12a) kullanabilir: biri bir AD ile kaynaşmak için, diğeri çıplak veya bir baskı alanı ile kombinasyon halinde. Karmaşıklık puanı yalnızca bir birim artacaktır (S = 12). Bu nedenle, CRISPR-dCas sistemleri, S. cerevisiae’de çok karmaşık gen dijital devrelerinin inşasının anahtarıdır.
Bu makale, mayadaki hem dCas9- hem de dCas12a bazlı baskılayıcıların ve aktivatörlerin verimliliğini derinden karakterize etmektedir. Sonuçlar, aktivitelerini optimize etmek için yüksek miktarda sgRNA talep etmediklerini göstermektedir, bu nedenle epizomal plazmidlerden tercihen kaçınılmaktadır. Dahası, dCas9 tabanlı aktivatörler, VP64 AD’nin kopyalarını toplayan bir iskele RNA’sı (scRNA) kullanıldığında çok daha etkilidir. Buna karşılık, dCas12a doğrudan güçlü VPR AD ile kaynaştırıldığında iyi çalışır. Ayrıca, sentetik olarak aktive edilmiş bir promotör, aktivatörün konfigürasyonuna bağlı olarak değişken sayıda hedef bölge gerektirir (örneğin, dCas12a-VPR kullanırken üç, dCas9-VP64 için altı ve dCas9 ve scRNA ile sadece bir tane). Bir baskılayıcı olarak, dCas12a, promotörden ziyade kodlama bölgesini bağlarken daha keskin görünür.
Bununla birlikte, bir dezavantaj olarak, CRISPR-dCas9 / dCas12a kimyasallarla doğrudan etkileşime girmez. Bu nedenle, giriş katmanında hiçbir işe yaramayabilirler. Bu nedenle, anti-CRISPR proteinleri (Acrs) içeren alternatif Boole kapısı tasarımları araştırılmıştır. Akr’ler (d)Cas proteinleri üzerinde etkili olur ve çalışmalarını engeller8. Bu nedenle, CRISPR-(d)Cas sistemlerinin aktivitesini modüle etmenin bir yoludur. Bu yazıda S. cerevisiae’de tip II Acrs ve (d)Cas9 ile tip V Acrs ve (d)Cas12a arasındaki etkileşimler kapsamlı bir şekilde analiz edilmiştir. Acrs, Cas proteinlerinden çok daha küçük olduğundan, östrojen β-östradiol’e yanıt veren bir NOT kapısı, insan östrojen reseptörü 9-HBD’nin (hER) hormon bağlama alanını AcrIIA4’e kaynaştırarak inşa edilmiştir. Ayrıca, dCas12a(-AD)’yi yapısal olarak ifade eden bir avuç EVET ve DEĞİL kapısı ve galaktoz ile indüksiyon üzerine AcrVA’lar gerçekleştirildi. Şu anda, bu kapılar sadece bir kavram kanıtı olarak hizmet vermektedir. Bununla birlikte, aynı zamanda, maya hücrelerindeki sentetik gen dijital devrelerinin hesaplamalı otomatik tasarımını gerçekleştirmek için algoritmanın derinlemesine yeniden düşünülmesine yönelik ilk adımı temsil ederler.
