Strakke barrières scheiden het centrale zenuwstelsel (CZS) van de periferie, inclusief de bloed-hersenbarrière (BBB) en de bloed-hersenvocht (CSF) barrière. Deze barrières beschermen het CZS tegen externe beledigingen en zorgen voor een evenwichtige en gecontroleerde micro-omgeving 1,2,3. Hoewel de BBB in de loop van de tijd uitgebreid is bestudeerd, heeft de bloed-CSF-barrière bij de choroïde plexus (CP) het afgelopen decennium alleen maar meer onderzoeksinteresse gekregen. Deze laatste barrière is te vinden in de vier ventrikels van de hersenen (figuur 1A, B) en wordt gekenmerkt door een enkele laag vaatvliesplexusepitheelcellen (CPE) rond een centraal stroma, lekkende haarvaten, fibroblasten en een lymfoïde en myeloïde celpopulatie (figuur 1C)4,5,6. De CPE-cellen zijn stevig met elkaar verbonden door tight junctions, waardoor lekkage van de onderliggende fenestrated bloedcapillairen naar de liquor en de hersenen wordt voorkomen. Bovendien wordt het transport over de CPE-cellen gereguleerd door een aantal in- en uitgaande transportsystemen die de instroom van nuttige verbindingen (bijv. Voedingsstoffen en hormonen) van het bloed naar de liquor en de efflux van schadelijke moleculen (bijv. Metabolisch afval, overtollige neurotransmitters) in de andere richting beheren 1,6. Om hun actieve transportfunctie te kunnen uitoefenen, bevatten de CPE-cellen talrijke mitochondriën in hun cytoplasma7. Bovendien is de CP de belangrijkste bron van CSF en fungeert als de poortwachter van de hersenen door de aanwezigheid van residente ontstekingscellen1. Door zijn unieke locatie tussen het bloed en de hersenen is de CP ook perfect gepositioneerd om immuunsurveillance uit te voeren8.
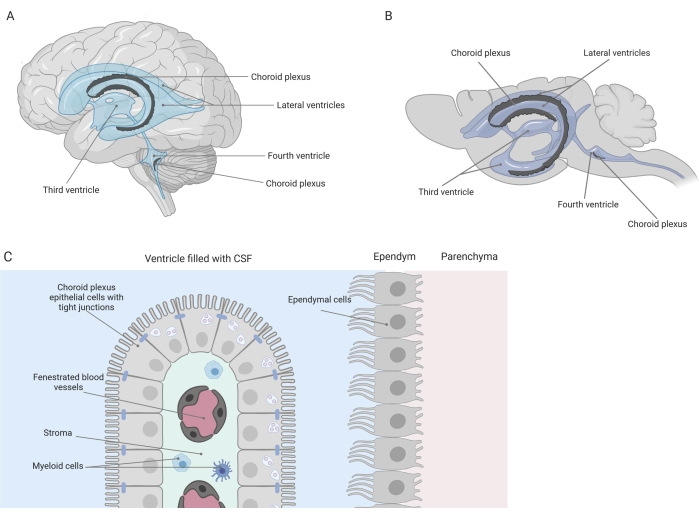
Figuur 1: Schematisch overzicht van de locatie en samenstelling van de plexus choroideus (CP). (A,B) CP-weefsel wordt gevonden in de twee laterale, de derde en de vierde ventrikels van (A) menselijke en (B) muizenhersenen. (C) Het CP-weefsel bestaat uit een enkele laag nauw verbonden kubusvormige CP-epitheel (CPE) -cellen rond fenestratieve haarvaten, los bindweefsel en lymfoïde en myeloïde cellen, en vormt de bloed-cerebrospinale vloeistofbarrière (aangepast en gewijzigd op basis van referentie23). Figuur gemaakt met Biorender.com. Klik hier om een grotere versie van deze figuur te bekijken.
In het afgelopen decennium hebben toenemende bewijzen, waaronder verschillende rapporten van onze onderzoeksgroep, aangetoond dat de CP een centrale rol speelt in gezondheid en ziekte 9,10,11,12,13,14,15,16,17,18 . Zo is bekend dat de verouderende bloed-CSF-barrière morfologische veranderingen vertoont in onder andere de kernen, microvilli en het keldermembraan 1,19. Bovendien is in de context van de ziekte van Alzheimer de algehele barrière-integriteit aangetast en al deze leeftijdsgerelateerde veranderingen lijken nog meer uitgesproken te zijn 1,8,20. Naast morfologische veranderingen zijn het transcriptoom, proteoom en secretoom van de CP veranderd tijdens ziekte 12,21,22,23. Geavanceerde kennis van de CP is dus essentieel om de rol ervan bij neurologische ziekten beter te begrijpen en mogelijk nieuwe therapeutische strategieën te ontwikkelen.
Een efficiënte methode voor nauwkeurige microdissectie van de CP uit de hersenventrikels is de eerste onschatbare stap om goed onderzoek van deze kleine hersenstructuur mogelijk te maken. Vanwege zijn sterk gevasculariseerde aard (figuur 2B) kan de CP die in de ventriculaire holtes van de hersenen zweeft, worden geïdentificeerd met behulp van een binoculaire microscoop. Transcardiale perfusie is echter vaak vereist voor downstream-analyse, wat de juiste identificatie en isolatie van het CP-weefsel bemoeilijkt (figuur 2C). Als de verdere verwerkingsstappen het toelaten (bijvoorbeeld in het geval van RNA- en eiwitanalyse), kan de CP worden gevisualiseerd via transcardiale perfusie met broomfenolblauw (figuur 2A). Verschillende publicaties beschrijven al de isolatie van de CP van rat24 en muizenpuphersenen25. Hier wordt een microdissectie-isolatietechniek beschreven om de CP te isoleren van volwassen muizen. Belangrijk is dat deze isolatietechniek de levensvatbaarheid, functie en structuur van de cellen binnen de CP behoudt. De isolatie van de CP zwevend in de vierde en laterale ventrikels wordt hier beschreven. Kortom, de muizen worden terminaal verdoofd en indien nodig transcardiaal geperfuseerd. Er moet echter worden opgemerkt dat perfusie de structuur van de cellen in de CP kan beschadigen. Als het monster moet worden geanalyseerd met behulp van transmissie-elektronenmicroscopie (TEM), seriële blokgezichtscanning elektronenmicroscopie (SBF-SEM) of gefocusseerde ionenbundel SEM (FIB-SEM), mag perfusie dus niet worden uitgevoerd. Vervolgens worden de hele hersenen geïsoleerd en wordt een tang gebruikt om de hersenen sagittaal te hemisecteren. Vanaf hier kunnen de CP’s die in de laterale ventrikels zweven worden geïdentificeerd en ontleed, terwijl de CP van de vierde ventrikel kan worden geïsoleerd van de cerebellaire kant van de hersenen.
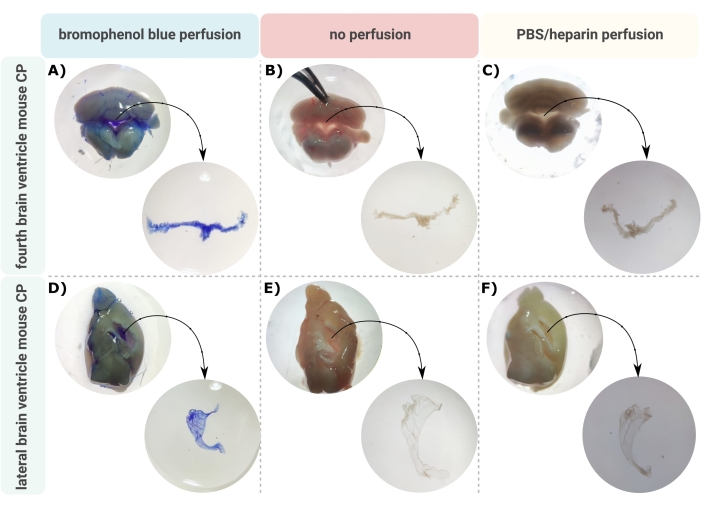
Figuur 2: Visualisatie van de (A-C) vierde en (D-F) laterale ventrikel choroïde plexus (CP) na (A,D) broomfenolblauwe perfusie, (B,E) geen perfusie en (C,F) perfusie met PBS/heparine. De beelden zijn gemaakt met een stereomicroscoop (8x-32x vergroting). Klik hier om een grotere versie van deze figuur te bekijken.
Zodra de CP goed is ontleed uit de hersenventrikels, kan een heel repertoire aan technieken worden toegepast om meer inzicht te krijgen in de functie van deze structuur. Flowcytometrie of eencellige RNA-sequencing kan bijvoorbeeld worden uitgevoerd om de infiltrerende ontstekingscellen onder bepaalde ziekteomstandigheden te kwantificeren en fenotypisch te analyseren26,27. Naast de cellulaire samenstelling kan de moleculaire samenstelling van de CP worden geanalyseerd om de aanwezigheid van cytokines en chemokines te beoordelen via enzyme-linked immunosorbent assay (ELISA), immunoblot of door gelijktijdige analyse van meerdere cytokines met behulp van de cytokine bead array28. Bovendien kunnen transcriptoom-, vasculaire, immuuncelhistologie en secretoomanalyses worden uitgevoerd op de microdissected CP-explantaten29. Hier wordt scanning elektronenmicroscopie (SEM) op whole mount CP gebruikt om een totaalbeeld van de CP-structuur te verkrijgen. SEM gebruikt een bundel van gefocuste elektronen om over het oppervlak te scannen en een beeld te creëren van de topografie en samenstelling van het oppervlak. Omdat de golflengte van elektronen veel kleiner is dan die van licht, ligt de resolutie van SEM in het nanometerbereik en superieur aan die van een lichtmicroscoop. Bijgevolg kunnen morfologische studies op subcellulair niveau worden uitgevoerd via SEM. Kortom, de ontleedde CP wordt onmiddellijk overgebracht in een glutaaraldehyde-bevattend fixatief voor een nachtelijke fixatie, gevolgd door osmicatie en uranylacetaatkleuring. De monsters worden vervolgens behandeld met loodaspartaatvlek, gedehydrateerd en uiteindelijk ingebed voor beeldvorming.
Dit protocol vergemakkelijkt dus de efficiënte isolatie van de CP van de hersenventrikels van de muis, die verder kunnen worden geanalyseerd met behulp van een verscheidenheid aan downstream-technieken om de structuur en functie ervan te onderzoeken.
