Des barrières serrées séparent le système nerveux central (SNC) de la périphérie, y compris la barrière hémato-encéphalique (BHE) et la barrière hémato-céphalo-rachidien (LCR). Ces barrières protègent le SNC contre les agressions extérieures et assurent un microenvironnement équilibré et contrôlé 1,2,3. Alors que la BHE a été largement étudiée au fil du temps, la barrière hémato-LCR située au plexus choroïde (PC) n’a fait que susciter un intérêt croissant de la recherche au cours de la dernière décennie. Cette dernière barrière se trouve dans les quatre ventricules du cerveau (Figure 1A, B) et se caractérise par une seule couche de cellules épithéliales du plexus choroïde (CPE) entourant un stroma central, des capillaires qui fuient, des fibroblastes et une population de cellules lymphoïdes et myéloïdes (Figure 1C)4,5,6. Les cellules CPE sont fermement interconnectées par des jonctions serrées, empêchant ainsi les fuites des capillaires sanguins fenêtrés sous-jacents dans le LCR et le cerveau. De plus, le transport à travers les cellules CPE est régulé par un certain nombre de systèmes de transport vers l’intérieur et vers l’extérieur qui gèrent l’afflux de composés bénéfiques (p. ex. nutriments et hormones) du sang vers le LCR et l’efflux de molécules nocives (p. ex. déchets métaboliques, neurotransmetteurs en excès) dans l’autre sens 1,6. Pour pouvoir exercer leur fonction de transport actif, les cellules CPE contiennent de nombreuses mitochondries dans leur cytoplasme7. De plus, le CP est la principale source de LCR et agit comme le gardien du cerveau par la présence de cellules inflammatoires résidentes1. En raison de son emplacement unique entre le sang et le cerveau, le PC est également parfaitement positionné pour effectuer une surveillance immunitaire8.
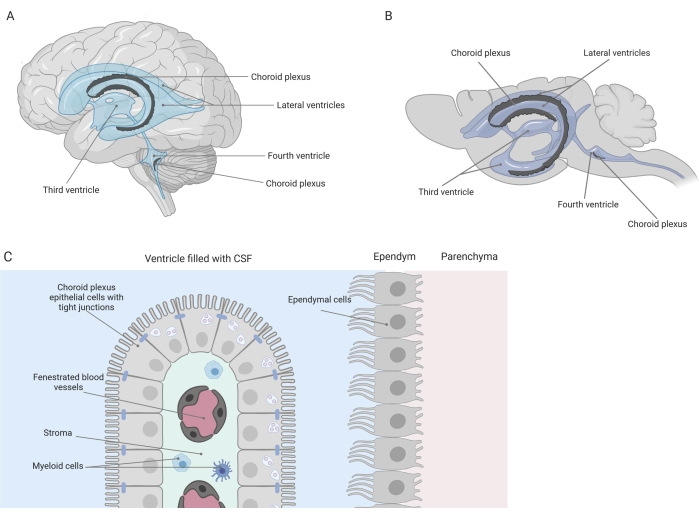
Figure 1 : Vue d’ensemble schématique de l’emplacement et de la composition du plexus choroïde (CP). (A,B) Le tissu CP se trouve dans les deux ventricules latéraux, le troisième et le quatrième ventricules (A) du cerveau humain et (B) du cerveau de la souris. (C) Le tissu CP est constitué d’une seule couche de cellules cuboïdes de l’épithélium CP (CPE) étroitement connectées entourant les capillaires fenêtrés, le tissu conjonctif lâche et les cellules lymphoïdes et myéloïdes, et forme la barrière hémato-céphalorachidienne (adaptée et modifiée à partir de la référence23). Figure créée avec Biorender.com. Veuillez cliquer ici pour voir une version agrandie de cette figure.
Au cours de la dernière décennie, de plus en plus de preuves, y compris plusieurs rapports de notre groupe de recherche, ont révélé que la PC joue un rôle central dans la santé et la maladie 9,10,11,12,13,14,15,16,17,18 . Par exemple, on sait que la barrière hémato-LCR vieillissante présente des altérations morphologiques dans, entre autres, les noyaux, les microvillosités et la membrane basale 1,19. De plus, dans le contexte de la maladie d’Alzheimer, l’intégrité globale de la barrière est compromise et tous ces changements liés à l’âge semblent être encore plus prononcés 1,8,20. En plus des changements morphologiques, le transcriptome, le protéome et le sécrétome de la PC sont modifiés au cours de la maladie 12,21,22,23. Ainsi, une connaissance avancée de la PC est essentielle pour mieux comprendre son rôle dans les maladies neurologiques et potentiellement développer de nouvelles stratégies thérapeutiques.
Une méthode efficace de microdissection précise de la PC hors des ventricules cérébraux est la première étape inestimable pour permettre une étude appropriée de cette minuscule structure cérébrale. En raison de sa nature hautement vascularisée (Figure 2B), la PC flottant à l’intérieur des cavités ventriculaires du cerveau peut être identifiée à l’aide d’un microscope binoculaire. Cependant, la perfusion transcardique est souvent nécessaire pour l’analyse en aval, ce qui complique l’identification et l’isolement corrects du tissu CP (Figure 2C). Si les étapes de traitement ultérieures le permettent (par exemple, dans le cas de l’analyse de l’ARN et des protéines), la PC peut être visualisée par perfusion transcardique avec du bleu de bromophénol (Figure 2A). Plusieurs publications décrivent déjà l’isolement du PC à partir de cerveaux de rats24 et de petits souris25. Ici, une technique d’isolement par microdissection est décrite pour isoler la PC des souris adultes. Il est important de noter que cette technique d’isolement préserve la viabilité, la fonction et la structure des cellules au sein du CP. L’isolement de la PC flottant dans le quatrième ventricule et les ventricules latéraux est décrit ici. En bref, les souris sont anesthésiées en phase terminale et, si nécessaire, perfusées par voie transcardale. Cependant, il convient de noter que la perfusion peut endommager la structure des cellules au sein du CP. Par conséquent, si l’échantillon doit être analysé par microscopie électronique à transmission (MET), microscopie électronique à balayage de face de bloc série (SBF-SEM) ou MEB à faisceau d’ions focalisés (FIB-MEB), la perfusion ne doit pas être effectuée. Ensuite, tout le cerveau est isolé et des forceps sont utilisés pour hémisecter le cerveau de manière sagittique. De là, les PC flottant dans les ventricules latéraux peuvent être identifiés et disséqués, tandis que les PC du quatrième ventricule peuvent être isolés du côté cérébelleux du cerveau.
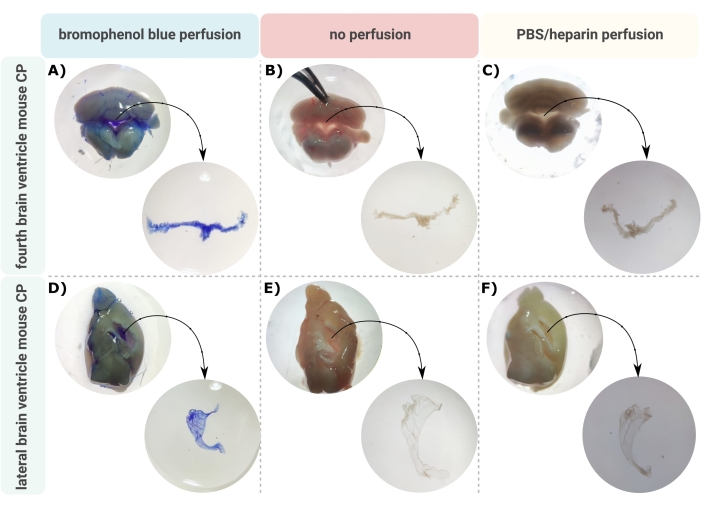
Figure 2 : Visualisation du (A-C) quatrième et (D-F) ventricule latéral du plexus choroïde (CP) après perfusion au bleu de bromophénol (A,D), (B,E) pas de perfusion et (C,F) perfusion avec PBS/héparine. Les images sont prises au stéréomicroscope (grossissement 8x-32x). Veuillez cliquer ici pour voir une version agrandie de cette figure.
Une fois que la PC est correctement disséquée hors des ventricules cérébraux, tout un répertoire de techniques peut être appliqué pour mieux comprendre la fonction de cette structure. Par exemple, la cytométrie en flux ou le séquençage de l’ARN unicellulaire peuvent être effectués pour quantifier et analyser phénotypiquement les cellules inflammatoires infiltrantes dans certaines conditions pathologiques26,27. En plus de la composition cellulaire, la composition moléculaire du PC peut être analysée pour évaluer la présence de cytokines et de chimiokines via un test immuno-enzymatique (ELISA), immunoblot, ou par analyse simultanée de plusieurs cytokines à l’aide du réseau de billes de cytokines28. De plus, des analyses du transcriptome, vasculaire, histologie des cellules immunitaires et sécrétome peuvent être effectuées sur les explants CP microdisséqués29. Ici, la microscopie électronique à balayage (MEB) sur l’ensemble de la monture CP est utilisée pour obtenir une vue d’ensemble de la structure de la CP. Le MEB utilise un faisceau d’électrons focalisés pour balayer la surface et créer une image de la topographie et de la composition de la surface. Comme la longueur d’onde des électrons est beaucoup plus petite que celle de la lumière, la résolution du MEB est de l’ordre du nanomètre et supérieure à celle d’un microscope optique. Par conséquent, les études morphologiques au niveau subcellulaire peuvent être effectuées via SEM. En bref, le CP disséqué est immédiatement transféré dans un fixateur contenant du glutaraldéhyde pour une fixation nocturne, suivie d’une osmication et d’une coloration à l’acétate d’uranyle. Les échantillons sont ensuite traités avec une coloration à l’aspartate de plomb, déshydratés et finalement intégrés pour l’imagerie.
Ainsi, ce protocole facilite l’isolement efficace de la PC des ventricules cérébraux de la souris, qui peut être analysée plus en détail à l’aide d’une variété de techniques en aval pour étudier sa structure et sa fonction.
