The pressure and current outputs of the respirometer controller are shown for one chamber in one experiment in Figure 3A. The first, long current pulse pressurized the chamber from ambient pressure (approximately 992 hPa) to the pre-set OFF threshold of 1017 hPa. As the flies consumed O2 and CO2 was absorbed, pressure decreased slowly until it reached the ON threshold of 1016 hPa, which activated current through the O2 generator. In the example shown, the average amplitude of each pulse is 50.1 mA, the duration is 16.1 s, yielding a charge transfer of about 0.81 coulombs (C) per pulse. The total charge transfer for this chamber was 3.28 C over a total time of 240.0 min. Using the calculations described in Procedures with the mass and number of flies (23 flies weighing 14.9 mg total), O2 consumption for the group in this chamber was 3.19 mL h-1 g-1 or 2.07 µL h-1 fly-1.
The equipment can be set up easily, with a minimum of training, and performs reliably for many cycles of assembly and shutdown. Nonetheless, equipment must be maintained and inspected regularly, and experimental conditions must be controlled carefully. For example, the loss of a gastight seal, due to failure of a joint or stopcock, can lead to rapid pressurization cycles and spuriously high VO2 (Figure 3B). Additionally, temperature and humidity must remain stable inside the chamber. If temperature or humidity decreases, the resulting pressure drop will be interpreted erroneously as a O2 being consumed. Conversely, upward drift in temperature or humidity will counteract the pressure decrease caused by O2 consumption, and artificially reduce or eliminate the VO2 signal (Figure 3C).
The method was used to test VO2 of CASKΔ18 mutants, which were generated by imprecise excision of a transposable element from the CASK locus14, and in which locomotion is drastically reduced14,16. In wild-type w(ex33) controls, generated by precise excision of the transposable element, average mass-specific O2 consumption was 3.65 ± 0.24 mL·g-1·h-1 (n = 16 chambers; Figure 4A).
Despite their visibly reduced locomotion, CASKΔ18 mutants' VO2 was slightly but not significantly lower than that of controls (mean ± s.e.m.= 3.23 ± 0.13 mL·g-1·h-1; n = 11 groups; P = 0.08 Mann-Whitney u-test).
Because the validity of expressing metabolic rate in terms of body mass has been questioned18, O2 consumption was also analyzed on a per-fly basis (Figure 4B). Using this analysis, VO2 was significantly reduced in CASKΔ18 compared to wild-type controls (ex33: 2.22 ± 0.13 µL·fly-1·h-1; CASKΔ18: 1.58 ± 0.10 µL·fly-1·h-1; P = 0.0003, Mann-Whitney u-test). However, the mean mass of CASKΔ18 flies was >20% lower than that of ex33 controls (Figure 4C; ex33 0.61 ± 0.01 mg; CASKΔ18 0.51± 0.02 mg; P = 0.0005, Mann-Whitney u-test), so the difference in metabolic rate between genotypes is probably due to the difference in their sizes.

Figure 1: Respirometer setup. (A) Diagram of sensor plug (above) and 50 mL chamber (consisting of a 50 mL Schlenk tube, below) before assembly. Note the locations of the 19/22 ground glass joints that will connect the chamber and sensor plug, and that must be cleaned before each experiment. The stopcock, which is necessary for opening or sealing the chamber, is also indicated. (B) Diagram of chamber and components, assembled and ready for the experiment, showing: wet cotton roll, polypropylene tube containing flies, plugged with a cotton stopper, soda-lime cartridge, and O2 generator filled with CuSO4. (C) Photograph of the assembled chamber. The Keck clamp securing the plug to the chamber is partially obscured by the ring stand clamp holding the chamber. (D) Photograph of controller showing switch controlling current through O2 generator and window for viewing OLED display. (E) Assembled chambers in a water bath. Seven chambers are shown, with three containing mutants, three with wildtype controls, and one chamber containing all components except flies. Please click here to view a larger version of this figure.
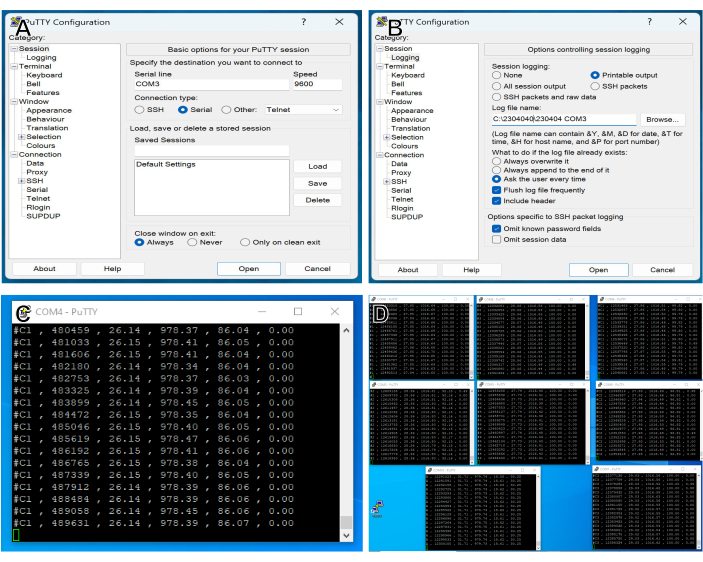
Figure 2: Data acquisition setup. (A) PuTTY interface for selecting the serial port for data acquisition. COM3 has been selected, with a BAUD rate of 9600 to match the output of the controller. (B) PuTTY interface for setting up log file. "Printable output" is selected to enable logging of data to a text file, data folder is selected using the "Browse" button, and a filename is created. (C) PuTTY log file during an experiment. Data are acquired approximately twice per second, and each line contains the following comma-delimited information: Sensor Number, time (ms) since the beginning of acquisition, chamber temperature (°C), chamber pressure (hPa), Humidity (percent relative), and current (mA). (D) Data logging during a typical experiment, with seven windows for experimental chambers, plus one channel recording bath temperature, ambient air temperature, pressure, and humidity. Please click here to view a larger version of this figure.
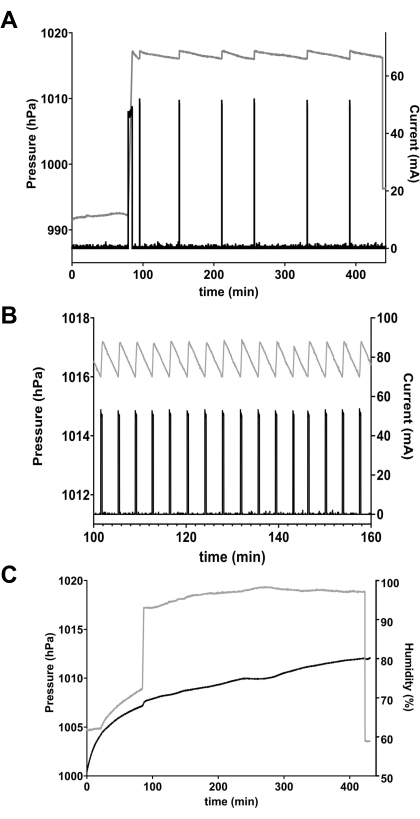
Figure 3: Data from microrespirometer. (A) Pressure (grey line, left axis) and current across the O2 generator (black line, right axis) in a single respirometer chamber containing 23 w(ex33) flies. At the beginning, a long current pulse is required to pressurize the chamber from ~992 hPa to the OFF threshold of 1017 hPa. As the flies consumed O2, pressure dropped until it reached the ON threshold of 1016 hPa, which activated current through the O2 generator, which re-pressurized the chamber to 1017 hPa. The process was repeated six times in this experiment. (B) An example of a leaky chamber caused by a damaged stopcock, taken from a different series of experiments. The chamber failed to maintain pressure (grey line), resulting in constant cycling of electrolytic current (black line). Note different timescale from panel A. (C) Effect of drift in humidity. O2 consumption by the 20 mg lady beetle (Hippodamia convergens) in the chamber should have produced a cycling pattern of pressure similar to Figure 3A, but the steady increase in humidity (black line) caused an artifactual increase in chamber pressure (grey line) that masked VO2. Please click here to view a larger version of this figure.
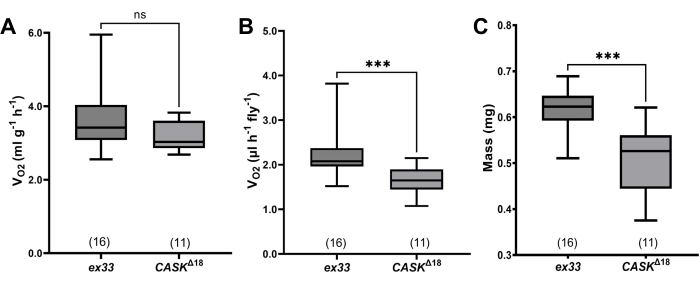
Figure 4: Quantitative data from wild-type and CASK mutant D. melanogaster. (A) Mass-specific VO2 for wild-type (w(ex33)) and mutant (CASKΔ18) flies. In all plots, bottoms and tops of boxes indicate first and third quartiles, respectively, whiskers indicate extreme values, and the sample sizes (numbers of chambers, each containing 17-24 flies) are given in parentheses above the genotypes. CASKΔ18 flies are not statistically significantly different from ex33 (median: ex33: 3.420 mL·g-1·h-1, CASKΔ18: 3.029 mL·g-1·h-1; p = 0.08 Mann-Whitney u-test). (B) Fly-specific VO2. CASKΔ18 flies consumed significantly less O2 (median 1.650 mL·fly-1·h-1) than w(ex33) (2.078 mL·fly-1·h-1; p = 0.0003, Mann-Whitney u-test; significance indicated by asterisks). (C) Mass differed between CASKΔ18 and the wildtype (median: 0.526 mg, ex33: 0.623 mg; p = 0.0005, Mann-Whitney u-test; significance indicated by asterisks). Please click here to view a larger version of this figure.
Table 1. Survey of Drosophila respirometry data from wildtype flies at 25 °C. With one exception18, studies are limited to those measuring O2 consumption. In most cases, it was necessary to estimate VO2 from graphs, and figure numbers from the original papers are provided. Although all genotypes were considered to be "wildtype" the sources and propagation methods varied. Please click here to download this Table.
