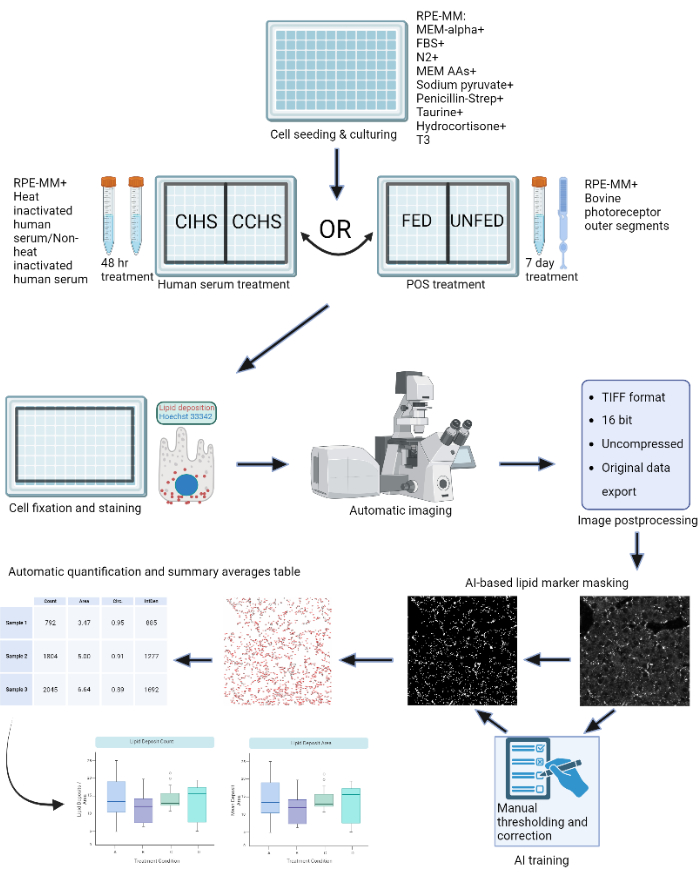
Alle protokolltrinn følger retningslinjene fastsatt av NIHs komité for human forskningsetikk. Stamcellearbeid og pasientprøveinnsamling ble godkjent av Combined NeuroScience Institutional Review Board (CNS IRB) under Office of Human Research Protection (OHRP), NIH, i henhold til 45 CFR 46 retningslinjer fra den amerikanske regjeringen. Pasientprøver ble samlet inn ved hjelp av CNS IRB-godkjent samtykkeskjema i henhold til kriteriene satt av Helsinkideklarasjonen under protokollnummer NCT01432847 (https://clinicaltrials.gov/ct2/show/NCT01432847?cond=NCT01432847&draw=2&rank=1). 1. iRPE generasjon Generer iRPE fra pasientblodavledet iPSC etter publisert protokoll av Sharma et al., 202211 (figur 2 og figur 3). Figur 2: Skjematisk fremstilling av iRPE-differensiering og modning. For å generere iRPE ble en etablert differensieringsprotokoll fulgt, og cellene fikk modnes i 5 uker. Den resulterende cellekulturen fungerer som en in vitro-modell som kan manipuleres med ulike behandlinger for å etterligne RPE-dysfunksjon i sykdommer som AMD og Stargardt sykdom. Klikk her for å se en større versjon av denne figuren. Figur 3: Representative bilder av vellykket og mislykket RPE-differensiering og modning. To lysfeltbilder ved 10x forstørrelse av TJP1 RPE vises på dag 42 av iRPE-protokollen. (A) Vellykket differensiering og modning vil vise konfluent RPE med pigmentering og polygonal morfologi. (B) Mislykket differensiering og modning vil vise klynger av døende celler, som vist her. Klikk her for å se en større versjon av denne figuren. 2. RPE vedlikeholdsmedier (RPE-MM) forberedelse Tine N2-tilskuddet ved 4 °C over natten. Tine alle andre reagenser ved romtemperatur (RT). Under sterile forhold, legg til reagensene oppført i tabell 1 ved de oppførte fortynningsfaktorene, i henhold til protokollen etablert av Sharma et al., 202211. Bland mediet godt og filtrer det med en 0,22 μm filtreringsenhet.MERK: Media er egnet for bruk innen 2 uker hvis de oppbevares ved 4 °C. 3. 96-brønns platesåing Tine en alikot av vitronectin ved RT i 3-5 min eller til isen er helt smeltet. Fortynn vitronectin med 1x Dulbeccos fosfatbufrede saltvann (DPBS) for å oppnå ønsket arbeidsløsning ved bruk av en 1:200 fortynning (vitronectin: DPBS). For en 96-brønnsplate, belegg hver brønn med 200 μL av arbeidsløsningen. Kombiner tint ROCK-hemmer (Y-27632 dihydroklorid) med RPE-MM ved en 1:1000 fortynning for å oppnå en endelig konsentrasjon på 10 μM. Dette er pletteringsmediet for RPE-celler. Tine iRPE-hetteglasset ved hjelp av et automatisert celletinesystem og overfør iRPE-cellesuspensjonen til en 50 ml slange med slanger. Fortynn cellesuspensjonen med pletteringsmediet ved en fortynning på 1:10. Sentrifuger røret ved 400 x g i 5 minutter. Aspirer supernatanten forsiktig og resuspender cellene i 10 ml av pletteringsmediet. Bland 400 mikrol av pletteringsmediet med 100 mikrol av den resuspenderte celleoppløsningen for celletelling. Bruk denne alikoten til å bestemme den levedyktige cellekonsentrasjonen av cellesuspensjonen ved hjelp av en cellelevedyktighetsteller. Fortynn cellesuspensjonen med pletteringsmediet til en endelig konsentrasjon på 60 000 celler/ml. Aspirer vitronectinbeleggløsningen helt fra 96-brønnplaten og dispenser 200 μL av cellesuspensjonen i hver brønn. Det vil være omtrent 12.000 celler / brønn eller ~ 200 celler / mm2. Inkuber de sådde celleplatene i 48 timer ved 37 °C og 5 % CO2. Etter 48 timer, bytt media til RPE-MM uten ROCK inhibitor supplement. Bytt media hver 2-3 dag i løpet av 5-ukers modningsperioden. 4. In vitro sykdomsmodeller Komplementær kompetent human serum (CC-HS) behandlingTin komplementkompetent serum ved 4 °C over natten. Forbered CC-HS og utfyll inkompetente humane serummedier (CI-HS).For å tilberede 5 % CC-HS-medier, bland det tinte komplementkompetente humane serumet med RPE-MM ved en fortynning på 1:20. Filtrer oppløsningen gjennom et 0,22 μm mediefilter før bruk. For å forberede 5 % komplementære medier for inkompetent humant serum (CI-HS), må du først varme inaktivere CC-HS i et vannbad på 57 °C i 30 minutter og deretter blandes med dyrkningsmediet ved en fortynning på 1:20. Filtrer oppløsningen gjennom et 0,22 μm mediefilter før bruk. Serum behandler cellene med 200 μL av enten 5% CC-HS eller 5% CI-HS media for en total inkubasjonstid på 48 timer, forfriskende media etter 24 timer. Vask cellene med 1x DPBS og fiks dem med 4% paraformaldehyd i 20 minutter ved RT. Vask en gang til med 1x DPBS og oppbevar prøvene ved 4 °C, nedsenket i 200 μL DPBS. VALGFRITT: Hvis ønskelig, lyse cellene fra platen for å vise bare sub-RPE lipidavsetning.For å lyse cellene og la bare lipidavsetninger, fjern mediet og tilsett 200 μL avionisert vann til hver brønn. Inkuber i 10-15 min, pipett opp og ned til cellene er fjernet. Vask igjen med 200 μL avionisert vann og fest umiddelbart cellene med 4% paraformaldehyd. Bekreft cellefjerningseffekten med kjernefysisk farging ved hjelp av Hoechst. Tilsett Hoechst ved 1:2000 fortynning til en 1x DPBS-løsning inneholdende 1% bovint serumalbumin (BSA), 0,5% Tween 20 og 0,5% Triton-X 100. Inkuber ved RT i 1 time i mørket. Deretter vasker du med 1x DPBS. Behandling av fotoreseptor ytre segment (POS) på iRPEPOS-forberedelseFjern POS-pelletsrøret fra -80 °C og tine over natten ved 4 °C i en tildekket isbøtte. Forbered POS-vaskebuffer ved å blande 10 g sukrose i 40 ml dobbeltavionisertH2O(ddH2O). Varm blandingen ved 40-50 °C under omrøring forsiktig i 15 minutter. Tilsett 840 mg natriumbikarbonat til blandingen og rør under oppvarming i 10 minutter. Juster det totale volumet av POS-vaskebufferen til 100 ml med ddH2O og juster pH i løsningen til 8,3 med 1 N HCl eller 1 N NaOH, etter behov. Filtrer vaskeoppløsningen med et 0,22 μm filter.MERK: Protokollen kan settes på pause her; POS-vaskebufferen kan lagres ved 4 °C over natten. Når den er tint, suspender pelleten i 15 ml POS vaskebuffer. Vær forsiktig under pelletssuspensjoner for å sikre POS-integritet. Sentrifuger POS-suspensjonen ved 600 x g ved 4 °C i 20 minutter og aspirer deretter supernatanten. Suspender POS-pelleten på nytt i 10 ml av POS-vaskebufferen. Fjern en 100 μL alikot av POS + POS-vaskebufferen (POS-løsning) og fortynn i 400 μL 1x DPBS. Spred 50 μL av den fortynnede POS-løsningen på en blodagarplate og en agaroseplate for å kontrollere for bakterielle og soppforurensninger. Forbered positive kontroller for hver og inkuber alle platene i 48 timer ved 37 °C. Utfør en qPCR-analyse ved å legge til 1 μL POS-løsning til deteksjonsbrønnen for å teste for mykoplasma. For å amplifisere DNA-fragmenter, utfør 40 sykluser med denaturering (95 ° C, 15 s), og glødning og forlengelse (60 ° C, 1 min). De fremre og bakre primere for påvisning av mykoplasma i POS-prøven er som følger:Forward primer: GGA TTA GAT ACC CTG GTA GTC CAC GOmvendt primer: CGT CAA TTC CTT TAA GTT TCA CTC TTG GC Mål POS-konsentrasjon ved hjelp av en celleanalysator og aliquot etter behov. For en brønn med 96 brønnplater med RPE-celler er 3 x 106 POS tilstrekkelig. Det ønskede forholdet er 10 POS / RPE-celle. Oppbevar alikotene ved 80 °C for fremtidig bruk. Tillegg av POS til cellerTine hetteglass med POS i et isbad. Bland den beregnede mengden forberedt POS med RPE-MM og behandle cellene med POS en gang daglig i 7 dager.NOTAT: Forbered POS-løsningen fersk daglig. Vask cellene med 1x DPBS og fikser dem deretter med 4% paraformaldehyd i 20 minutter ved RT. Vask med DPBS en gang til og oppbevar prøver ved 4 °C, nedsenket i 200 μL DPBS. 5. Farging for sub-RPE innskudd Nilen rød farget protokollEtter PFA-fiksering, vask prøvene 3 ganger med 1x DPBS.MERK: Hvis den ikke brukes umiddelbart, kan protokollen settes på pause her, men prøver må lagres i en 1x DPBS + 0,02% natriumazidoppløsning ved 4 °C. For å fremstille Nilens røde stamløsning, oppløs Nilrødt pulver i aceton ved 3 mg/ml konsentrasjon. Inkuber i 15 min ved RT med periodisk blanding. Filtrer løsningene med et 0,22 μm filter en eller to ganger, avhengig av nivået av bunnfall som er igjen i løsningen.MERK: Beskytt stamløsningen mot lys. For å forberede arbeidsløsningen, fortynn stamoppløsningen i forholdet 1:500 i 1x DPBS. Legg 200 μL av arbeidsløsningen til prøven i 30 minutter ved RT på en shaker og beskytt den mot lys. Vask 3 ganger med 1x PBS og oppbevar prøvene ved 4 °C, nedsenket i 200 μl DPBS.MERK: Hvis du utfører et eksperiment på transbrønner i stedet for en 96-brønnsplate, kan prøvene monteres på et lysbilde med monteringsmedier, dekkes med et glassdeksel og forsegles med gjennomsiktig neglelakk. Det må utvises forsiktighet for å montere prøven med cellene vendt opp. BODIPY farging protokollFor stamløsning oppløses BODIPY i vannfritt dimetylsulfoksid (DMSO) for å oppnå en stamkonsentrasjon på 3,8 mM. For PFA-faste prøver, fortynn BODIPY-lager på 1:300 i 1x DPBS. Tilsett 200 μL til celler og inkuber over natten på en rocker ved RT. Vask 3 ganger med 1x DPBS og oppbevar prøvene ved 4 °C, nedsenket i 200 μl DPBS.MERK: Hvis du utfører et eksperiment på transbrønner i stedet for en 96-brønnsplate, kan prøvene monteres på et lysbilde med monteringsmedier, dekkes med et glassdeksel og forsegles med gjennomsiktig neglelakk. Det må utvises forsiktighet for å montere prøven med cellene vendt opp. APOE immunostaining protokollKombiner 1x DPBS med 1% bovint serumalbumin (BSA), 0,5% Tween 20 og 0,5% Triton-X 100 for å lage en bufferløsning. For PFA-faste prøver, blokker og permeabiliser prøven i 200 μL av bufferløsningen i 1 time ved RT. Tilsett APOE primært antistoff fortynnet ved 1:100 i bufferløsningen og inkuber over natten ved RT. Neste dag, vask prøvene 3 ganger med 1x DPBS. Legg til et sekundært antistoff ved 1:1000 fortynning i bufferløsningen og tilsett 200 μL av løsningen til cellene i 1 time ved RT. Vask 3 ganger med 1x DPBS og oppbevar prøver ved 4 °C, nedsenket i 200 μl DPBS.MERK: Hvis du utfører et eksperiment på transbrønner i stedet for en 96-brønnsplate, kan prøvene monteres på et lysbilde med monteringsmedier (Fluoromount), dekkes med et glassdeksel og forsegles med gjennomsiktig neglelakk. Det må utvises forsiktighet for å montere prøven med cellene vendt opp. 6. Bildeautomatisering og behandling Automatisert bildeskanningMERK: Zeiss LSM 800 invertert konfokal skanningsmikroskop og ZEN 3.2 (blå utgave) programvare ble brukt i denne studien. Forsikre deg om at 96-brønnsplaten varmes opp til RT i minst 60 minutter før avbildning for å unngå fokalplandrift under skanningen på grunn av endring i brytningsindeksen til mediet med temperaturendring.Ved hjelp av et konfokalmikroskop og et 40x objektiv, opprett en skanneprofil med passende fluorescerende kanaler for lipidmarkøren som brukes og eventuelle ekstra antistoffer. Bruk avmerkingsboksen Fliser til å konfigurere bildeautomatisering. For å kalibrere 96-brønnsplaten, sørg for at de riktige prøvebærermålingene legges inn og velges. Klikk deretter på Kalibrer-knappen for å kalibrere platen i henhold til instruksjonene, noe som krever bruk av 10x-målet. Velg visningen Avansert oppsett for å velge passende brønner og legge til 3 forskjellige bildepunkter nær midten av brønnen ved hjelp av funksjonen Posisjoner . Dette kan gjøres manuelt under underfanen Posisjon eller tilfeldig ved å bruke kategorien Posisjonsoppsett og velge Oppsett etter transportør. Gjenta for alle brønnene med samme farging. For optimal fokusering og Z-stack-posisjonering under automatisering, gå til kategorien Fokusstrategi for å velge Bruk fokusoverflate/Z-verdier definert av flisoppsett. Alternative metoder kan bruke andre fokusstrategier, men det anbefales å bruke denne innstillingen for å få de mest konsekvente resultatene. Klikk Kontroller posisjoner under kategorien Fliser, og angi det sentrale Z-planet manuelt for hver posisjon. Innstillingene i underfanen Alternativer vil bestille anskaffelse av bilder, så sjekk dette før du begynner å bilde. Hvis du vil hente bilder i den rekkefølgen posisjonene ble valgt, fjerner du merket for Flisområder/-posisjoner og Bærerbrønner/containere. Velg Del scener i separate filer for enkel bildebehandling. Kontroller at Z-Stack-fanen er satt til Midtstilt, at et område er lagt inn i brukerens innstillinger, og at Optimal-knappen er valgt for å angi stykkeintervallet. Når du har optimalisert fanene Anskaffelsesmodus, Kanaler, Fokusstrategi, Z-Stack og Fliser, starter du eksperimentet. BildebehandlingVed hjelp av en batchavbildningsmetode kan du lage maksimale projeksjoner av hver Z-stakk med metoden Utvidet dybdeskarphet . Bruk en batchbildebehandlingsmetode til å eksportere de maksimale projeksjonsfilene som 16-biters TIFF-bilder. Sett komprimeringen til Ingen og kontroller at det er merket av for Originale data . Det resulterende bildet skal være en maksimal projeksjonsgråskala TIFF av bare fluorescenskanalen som lipidmarkøren uttrykkes på. 7. Segmentering og kvantifisering MERK: LipidUNet-programmet ble trent på 40x bilder fra en 96-brønnsplate. Det anbefales sterkt å bruke bilder som ble oppnådd ved hjelp av et 40x-mål. Installer LipidUNet-programvaren. LipidUNet kan lastes ned fra følgende GitHub-depot: https://github.com/RPEGoogleMap/LipidUNet Identifiser TIFF-bildene som representerer enten Nile Red, Bodupy eller APOE, og flytt dem til en mappe som heter imgs i en katalog som heter enten Nile_Red, Bodipy, eller APOE, avhengig av metoden som brukes.MERK: Nøyaktige navnekonvensjoner må brukes for at LipidUNet-programmet skal gjenkjenne katalogene. Åpne LipidUNet-programvaren (figur 4). I kategorien Predict i programvaren velger du den aktuelle katalogen (Nile_Red, Bodipy , eller APOE) ved å klikke på ellipsen og navigere til den navngitte katalogen. Bekreft at LipidUNet-programmet har identifisert bildene riktig ved å sjekke klasseoppføringen. Velg en sannsynlighetsterskel for algoritmen mellom 0,01 og 0,99. En høyere verdi vil eliminere flere falske positiver, men kan føre til flere falske negativer, og lavere verdier kan introdusere flere falske positiver samtidig som flere falske negativer elimineres. En verdi på 0,65 er standard og anbefales. Klikk på Forutsi.MERK: Programvaren vil iterere gjennom alle bildene automatisk og opprette en ny mappe kalt predicted_masks i den valgte katalogen. Bruk et maskeanalyseverktøy til å gjenta gjennom de genererte maskene og gi et kvantitativt antall terskellipidavleiringer fra maskebildene. Analyser de genererte telledataene for å sammenligne behandlingsbetingelser. Figur 4: LipidUNet brukergrensesnitt. LipidUNet-programvaren har forskjellige seksjoner å velge for treningsdatakatalogen, der bilder av lipidavleiringer er identifisert riktig; modellvektkatalogen, som er produsert fra treningsdataene; og prediksjonsdatakatalogen der brukeren vil legge inn bildene sine for segmentering. Klikk her for å se en større versjon av denne figuren.