In this protocol, we used commercially produced probes targeting Mef2 and zfh1 mRNA. We designed the probes with the manufacturer's FISH probes designer (Table of Materials). The Mef2 set targets the exons common to all Mef2 RNA isoforms (from exon 3 to exon 10). The zfh1 set targets the third exon common to both zfh1-RB and zfh1-RE isoforms10. Both probes are conjugated to the Quasar 670 fluorescent dye.
We generated a macro in-house that was compatible with ImageJ software to automatically analyze the raw .tif data. The macro allows the 3D segmentation of the nuclei by Cellpose26 with the Fiji plugin BIOP and the transcriptional spot quantification by a Laplacian of Gaussian as implemented in the Trackmate plugin27. MorpholibJ plugin28 is used for postprocessing (Supplemental File 1).
As a first step, we performed smFISH on wing imaginal discs, where Mef2 and zfh1 are known to be expressed in the AMPs. These data show that both transcripts are uniformly detected in the AMPs population co-stained with either Mef2 or Zfh1 antibodies (Figure 2A,B). Active transcription sites (TS) can be revealed and distinguished from mature mRNA by smFISH as they tend to be larger and have more intense signals than individual cytoplasmic transcripts or mature nuclear transcripts. Consistently, higher magnifications of AMPs distinguish between TS foci and mature mRNA scattered in the cytoplasm (Figure 2A',B'), validating the sensitivity of this smFISH protocol. It should, however, be mentioned that we observed one active TS per nucleus for both zfh1 and Mef2 (Figure 2A',B'). These data further validate the accuracy of the probe design since Mef2 and zfh1 transcriptional patterns in the AMPs colocalize with the detection of their respective proteins (Figure 2).
In a second step, we examined the transcription site and distribution of Mef2 and zfh1 mRNAs in differentiated adult IFMs and associated stem cells (Figure 2C,D). These data clearly illustrate Mef2 TSs in the syncytial muscle nuclei and Mef2 mRNA distribution throughout the cytoplasm (Figure 2C,C'). Zfh1 specifically marks the MuSCs population10,11. Using this smFISH protocol, we successfully detected zfh1 transcription in MuSCs marked by Zfh1-Gal4 > GFP expression. Higher magnification illustrates the detection of both zfh1 TS and cytoplasmic single mRNAs (Figure 2D').
Finally, we used our in-house-built ImageJ macro to quantitatively analyze Mef2 transcriptional dynamics and spatial distribution. The computational pipeline can efficiently detect and segment Mef2 spots and muscle nuclei (Figure 3A–C). However, due to signal-to-noise ratio issues, some spots may not be detected in certain cases (Figure 3A',B'). We used this automated method to investigate whether Mef2 mRNA abundance varies between muscle precursor cells (AMPs) and differentiated adult IFMs. Our analysis shows that the number of Mef2 mRNA per nuclei in adult IFMs and the AMPs are not significantly different (Figure 3D).
To gain insight into the spatial distribution of Mef2 transcripts in adult IFM muscles, we quantified the fraction of cytoplasmic versus nuclear Mef2 mRNA spots (Figure 3E,F). Using our program, we counted both the total number of Mef2 mRNA spots and the number of Mef2 mRNA spots overlapping with nuclei staining (Mef2). Subtracting the nuclei-associated mRNA from the total mRNA number provides the number of cytoplasmic mRNA spots. Our findings reveal that approximately 92% of Mef2 mRNA is present in the cytoplasm, while the remaining 8% is associated with muscle nuclei, as shown in Figure 3E,F.
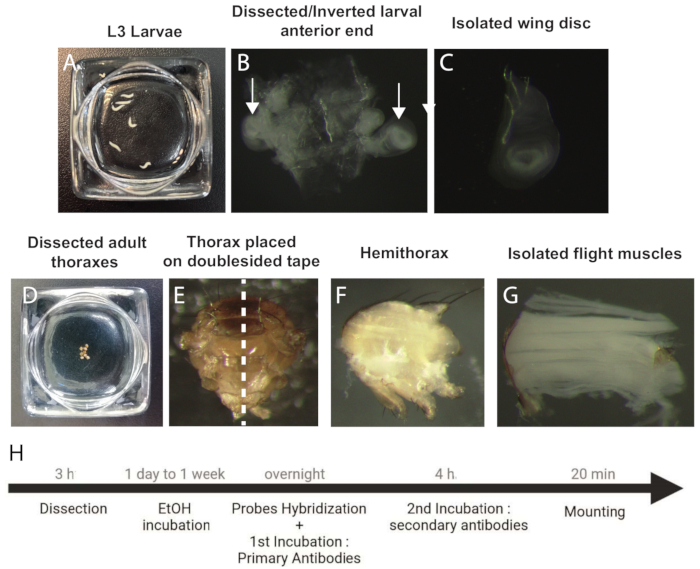
Figure 1: Procedure to dissect and prepare wing discs and IFM samples for smFISH. (A) L3 staged larvae. (B) Dissected and inverted anterior end of the larva. Arrows indicate the wing discs. (C) Isolated wing disc. (D) Dissected adult thoraxes. (E) Adult thorax oriented on a doublesided tape prior to bisection (dotted line). (F) Adult hemithorax. (G) Isolated IFMs. (H) Workflow of the smFISH protocol. Abbreviations: IFMs = indirect flight muscles; smFISH = single-molecule RNA fluorescence in situ hybridization; EtOH = ethanol. Please click here to view a larger version of this figure.
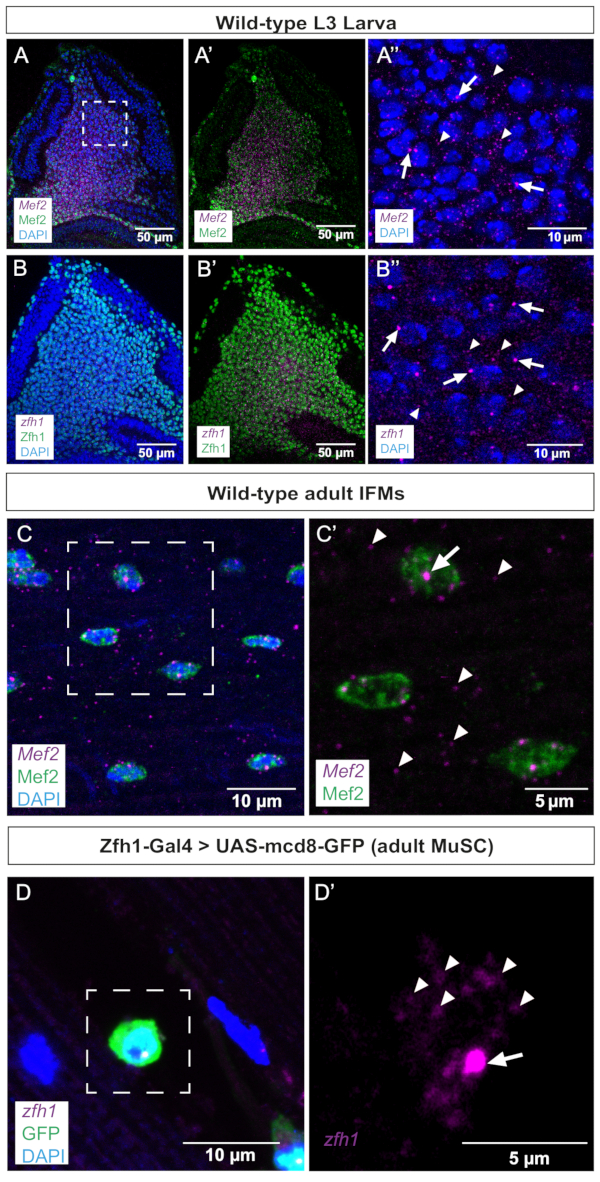
Figure 2: Representative images of Mef2 and zfh1 smFISH in wing discs and adult IFMs. (A,B) Mef2 and zfh1 mRNAs (purple) are detected uniformly in AMPs and co-localize with (A,A') Mef2 and (B,B') Zfh1 proteins (green), respectively. (A'',B'') Higher magnification of the AMPs. (C) Mef2 mRNAs and Mef2 protein are detected in adult IFMs. (C') Higher magnification of the boxed region in C. (D) Zfh1-Gal4 >UAS-mCD8GFP expression (green) labels adult MuSCs. zfh1 RNA is detected in adult MuSCs. (D') Higher magnification of the boxed region in D. Zfh1 transcription start sites and single mRNAs are indicated by arrows and arrowheads, respectively. DAPI staining (blue). Scale bars = 50 µm (A,B,A',B'), 10 µm (A",B",C,D), 5 µm (C',D'). Abbreviations: IFMs = indirect flight muscles; smFISH = single-molecule RNA fluorescence in situ hybridization; AMPs = adult muscle progenitors; MuSCs = muscle stem cells; DAPI = 4',6-diamidino-2-phenylindole; GFP = green fluorescent protein. Please click here to view a larger version of this figure.
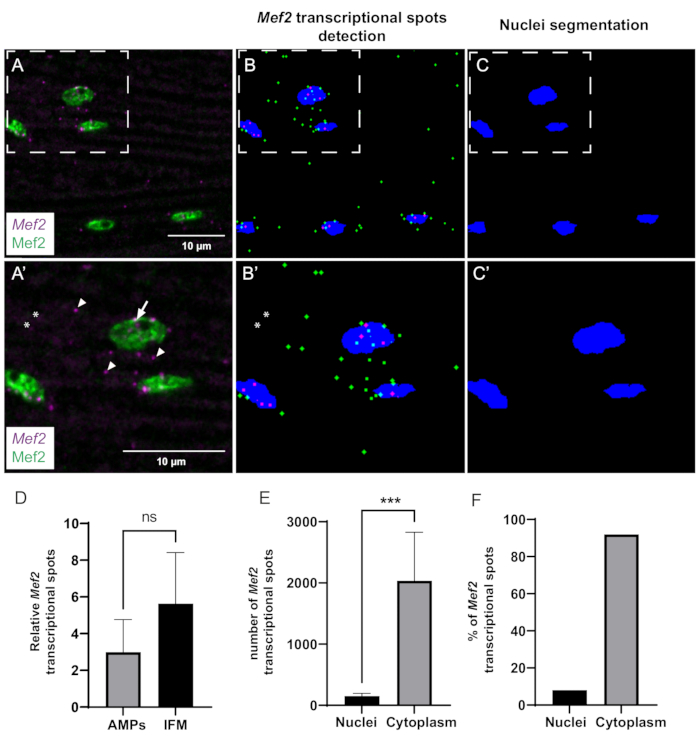
Figure 3: Quantification of Mef2 transcription and cytoplasmic mRNA accumulation by smFISH. (A) Distribution of Mef2 transcripts (purple) and Mef2 protein (green) in wildtype adult IFMs. (A') Higher magnification of boxed region in A. Transcription start sites and single mRNAs are indicated by arrows and arrowheads, respectively. (B,C) Automatic Mef2 spots finding and nuclear segmentation. Cytoplasmic and nuclear Mef2 mRNAs are indicated in green and magenta, respectively. (B',C') Higher magnifications of boxed regions in B and C. Asterisks indicate spots that are not counted by the macro. (D) Example of Mef2 spot quantification in AMPs and adult IFMs, relative to the total number of nuclei. (p = 0,0785, Student's t-test, wing discs (n = 4), IFMs (n = 11)). (E) Example of quantification of Mef2 spot distribution in adult IFMs. (*** p< 0.0007, Student's t-test, n = 5). (F) Percentage of Mef2 mRNA spots detected inside and outside the nuclei (n = 5). Scale bars = 10 µm (A,A'). Abbreviations: IFMs = indirect flight muscles; smFISH = single-molecule RNA fluorescence in situ hybridization; AMPs = adult muscle progenitors. Please click here to view a larger version of this figure.
Supplemental File 1: Macro used for postprocessing. Please click here to download this File.
