Cellemembranen, som fungerer både som en fysisk barriere og en signalplattform, er avgjørende for celleoverlevelse1. Gjennom hele cellesyklusen blir plasmamembranen (PM) utsatt for skader, for eksempel mekaniske 2,3,4,5 og kjemiske6 stressinduserte skader. For å opprettholde membranintegritet og sikre celleoverlevelse har cellen utviklet robuste mekanismer for reparasjon av plasmamembraner (PMR). Disse mekanismene avhenger av ulike strategier, for eksempel cytoskjelettomorganisering, membranfusjon og membranutskiftingsstrategier 7,8,9,10,11, som alle er avhengige av rekruttering av spesifikke proteiner. Spesielt har medlemmer av anneksinproteinfamilien blitt identifisert som nøkkelproteiner assosiert med prosessene til PMR 1,9,12,13,14,15,16. Etter PM-skade opplever cellen en tilstrømning av kalsiumioner (Ca2+), noe som utgjør en umiddelbar trussel mot cellens overlevelse17. Som svar på Ca2+ tilstrømning binder anneksinproteiner, som hovedsakelig er lokalisert i cytosolen, seg til den indre brosjyren til den skadede plasmamembranen som en del av PMR-strategiene18. Vedlegg A2 (ANXA2) var et av de første medlemmene av anneksinfamilien som ble assosiert med PMR ved dysferlinmangelfull muskeldystrofi og ble foreslått å formidle reparasjon ved å smelte intracellulære vesikler til PM nær skadestedet 5,19,20,21. I ettertid har flere funksjoner blitt tilskrevet vedlegg22, og deres rolle i PMR har fått økt oppmerksomhet de siste 20 årene. Den eksakte rollen til anneksiner i PMR er imidlertid ikke fullt ut forstått 15,18,21,22.
Denne artikkelen foreslår en metode for å undersøke proteinmembraninteraksjon og membrandynamikk på en kontrollert og svært lokalisert måte, ved hjelp av en kombinasjon av konfokalmikroskopi, optisk pinsett og gullnanopartikler (AuNPs). Denne metoden muliggjør kvantitativ studie av protein-, lipid- og småmolekylære interaksjoner som respons på membranskader og Ca2+ tilstrømning. Til tross for kompleksiteten og mangfoldet av komponenter som er involvert i prosessen med membranreparasjon, har forenklede membransystemer som etterligner plasmamembranen blitt brukt for å få en dypere mekanistisk forståelse av membrandynamikk og anneksinproteiners respons på membranforstyrrelser16. Giant unilamellar lipidvesikler (GUVer) ble valgt som modellmembransystem med en spesifisert lipidsammensetning. GUVene ble generert ved hjelp av gelassistert hydreringsmetode, spesielt polyvinylalkoholgelhydrering, som beskrevet av Weinberger et al.23, som tillot effektiv innkapsling av vedlegg i GUVer.
Utnyttelsen av nær-infrarød (NIR) laserbestråling på metalliske nanopartikler (NP) induserer betydelig oppvarming av NP, noe som gjør det til en effektiv metode for å etablere en lokal varmekilde utnyttet i biomedisinske applikasjoner24. Metoden ble opprinnelig brukt til å direkte måle temperaturen rundt en enkelt AuNP i både 2D og 3D biomimetiske analyser. I disse analysene 25,26 ble de plasmoniske nanopartiklene bestrålt på et støttet lipid-dobbeltlag eller optisk fanget nær GUVer som gjennomgikk en lokal termisk faseovergang ved lokal oppvarming, noe som muliggjorde kvantifisering og kontroll av den nøyaktige temperaturprofilen rundt partikkelen. Denne referansetemperaturprofilen har blitt brukt ved undersøkelse eller manipulering av biologiske prøver. Ytterligere fremskritt i metoden har gjort det lettere å induksjonere nanoskopiske porer i membraner27, noe som muliggjør vesikkel og cellefusjon 28,29. Andre studier har undersøkt oppførselen til perifere membranproteiner i GUVer29 og transmembranproteiner30 ved å skape nye hybridvesikler, mens cellespesifikk legemiddellevering også har blitt utforsket for å kontrollere og studere cellulære responser eller genuttrykk 28,29,31,32,33. Nylig har metoden blitt brukt til å undersøke proteinresponser på membranskader 32,34,35.
Det finnes flere metoder for å forstyrre plasmamembranen for å utforske cellulære responser og membranreparasjon. Disse inkluderer mikronålpunkteringer, mikroperleristing og celleskraping, som alle kan forstyrre cellemembranen mekanisk 14,36,37. Kjemisk indusert skade kan oppnås ved å tilsette vaskemidler 5,38 eller bakterielle toksiner39,40 som destabiliserer lipid-dobbeltlaget og genererer membranporer over plasmamembranen. Videre har optisk induserte skader ved kontinuerlige bølge- og pulserende lasere blitt brukt til å studere PMR-komponenter, slik som anneksinproteiner 5,14,21,41, i kombinasjon med plasmoniske nanopartikler 42,43,44,45 . Til tross for effektiviteten til pulserende lasere med høy effekt, kan de forårsake betydelige skader og skade på cellens indre langs strålebanen. Videre gjenstår de detaljerte endringene som skjer i det biologiske materialet ved pulserende laserbestråling og om det skaper en veldefinert pore, å bli undersøkt nærmere. En alternativ metode presenteres i denne artikkelen, ved bruk av termoplasmonikk for å indusere nanoskopiske hull i PM på en kontrollert måte34,35 uten å forårsake betydelig skade på de indre strukturer. Dette oppnås ved å utsette plasmoniske NP for en svært fokusert NIR-laser, noe som resulterer i en ekstremt lokalisert temperaturøkning som lett kan nå temperaturer over 200 ° C, noe som kan føre til små nanoskopiske eksplosjoner 25,46,47. Denne prosessen kan styres ved å justere laserintensiteten samt størrelsen, formen og sammensetningen av NPs48. Ved å bruke denne teknikken kan forskere utforske rollen som proteiner i PM-reparasjon i levende celler, noe som kan bidra til å løse noen av de ubesvarte spørsmålene om involvering av anneksinproteiner i membranreparasjon uten å kompromittere cellens levedyktighet.
Den optiske fangsten av plasmoniske nanopartikler har blitt godt etablert av tidligere studier 25,49,50,51,52; Imidlertid kan ytterligere innsikt om de termoplasmoniske egenskapene til nanopartiklene 53,54,55 fås i tilleggsmaterialene (tilleggsfil 1). Den termoplasmoniske metoden kan brukes til å lage nanoskopiske hull i PM med det formål å studere cellulær respons og reparasjonsmekanismer. Nærmere bestemt kan punkteringen oppnås ved optisk oppvarming av AuNP i nærheten av membranen, som vist i figur 1A og B. Denne lokaliserte punkteringen tillater Ca2+ tilstrømning, som ble verifisert av en kalsiumsensor, og dermed aktiverte PMR-maskineriet. For levende celleforsøk ble AuNP med en diameter på 200 nm immobilisert på overflaten under cellen for å overvåke ANXA2s rolle i PMR via konfokal mikroskopi. NIR-laseren (figur 1A,B), med en bølgelengde på 1064 nm, bestråler AuNP, utnytter dens plasmoniske egenskaper (figur 1C), noe som resulterer i betydelig lokal oppvarming (figur 1D) i det biologiske gjennomsiktighetsvinduet49 samtidig som den forårsaker minimal skade på selve cellen. Høytemperaturområdet rundt AuNP reduseres raskt med 30-40% i en avstand som tilsvarer NPs radius, som vist i figur 1E, noe som muliggjør en ekstremt begrenset skade i alle tre dimensjoner.
Tilleggsfil 1. Klikk her for å laste ned denne filen.
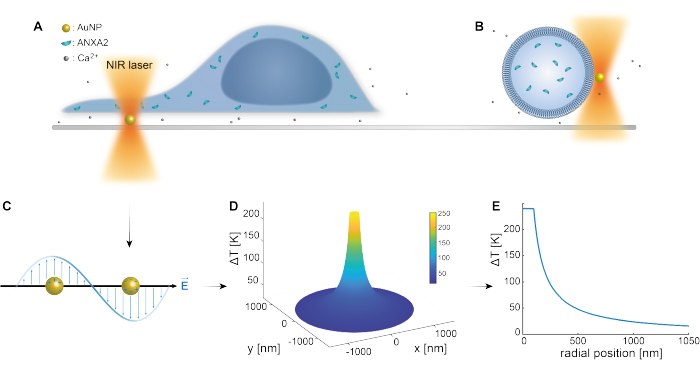
Figur 1: Skjematisk oversikt over eksperimentell metode. (A) ANXA-transfekterte celler ligger på toppen av immobiliserte gullnanopartikler (AuNPs) på overflaten, eller (B) gigantiske unilamellære vesikler (GUVer) med innkapslet ANXA er suspendert i et medium som inneholder AuNPs. (C) En enkelt AuNP bestråles av NIRs optiske felle, hvor vekselvirkningen mellom det innkommende elektromagnetiske feltet og ledningselektronene fører til den kollektive svingningen av elektroner i NP. (D) Denne prosessen resulterer i en svært begrenset, men likevel betydelig temperaturøkning. For å estimere temperaturen på NP-overflaten brukes Mie-teorien, og en (E) temperaturprofil beregnes for en AuNP med en diameter på 200 nm og laserintensitet I = 6,36 x 108 W/cm2. Klikk her for å se en større versjon av denne figuren.
For å minimere den termiske effekten på cellemembranen, blir AuNPene bare bestrålt i ~ 1 sekund. Dette fører til en forbigående og lokal utbrudd av oppvarming, noe som reduserer skaden på proteiner som vanligvis krever mer tid til å utfolde seg. Ved membranpunktering rekrutteres anneksinproteiner på brøkdelen av et sekund, og i løpet av få sekunder dannes et anneksinringlignende stillas rundt skadestedet (figur 2). Denne tilnærmingen har også blitt brukt til å utforske involvering av ANXA5 i både levende celler og modellmembraner16 i et forsøk på å kaste lys over hele skjemaet for reparasjonsprosessene. Mens hovedfokuset har vært på korrelerende rekruttering av ulike anneksinproteiner, har de biofysiske aspektene av reparasjonsmekanismen ennå ikke blitt belyst.
For å fullt ut implementere den foreslåtte metoden kreves tre nøkkelkomponenter: konfokalmikroskopi, optisk pinsett og metall nanopartikler. Optisk pinsett brukes til å fange AuNPs, og deres konstruksjon kan oppnås ved å følge prosedyren skissert av Neuman et al.49. Men hvis det viser seg å være for utfordrende å bygge en optisk pinsett, kan en tett fokusert NIR-laser brukes til å bestråle AuNPs immobilisert under cellene. Mens sfæriske AuNPer ble valgt for denne protokollen, kunne en rekke plasmoniske partikler med justerbare absorpsjonsspektra også benyttes for å oppnå en svært lokalisert temperaturgradient innenfor NIR-regionen48.
Fluorescensavbildning er nødvendig for å observere rollen til de fluorescerende merkede proteinene, og derfor kan total intern refleksjonsmikroskopi (TIRF)56 betraktes som et alternativ til konfokal avbildning. Denne teknikken tillater imidlertid bare overflateavbildning og vil ikke være kompatibel med modellmembranvesikelforsøkene. Følgelig er både den optiske pinsetten og konfokalmikroskopet avgjørende for den nøyaktige lokaliseringen av nanopartikkelen og detaljert undersøkelse av lokalområdet rundt celleskaden. For effektivt å bestråle nanopartikkelen med et diffraksjonsbegrenset laserfokus, er det nødvendig å visualisere nanopartikkelen. Dette kan oppnås optimalt ved refleksjonsmikroskopi, som er en standard avbildningsfunksjon i Leicas konfokale mikroskoper. Imidlertid, hvis refleksjon eller spredningsavbildning ikke er tilgjengelig, kan alternative metoder, for eksempel den mindre effektive fluorescerende AuNP-merkingen, vurderes.
Oppsummert har den svært kontrollerbare og lokaliserte termoplasmoniske metoden som presenteres i denne studien, potensial til å tjene som en utmerket plattform for å undersøke molekylære komponenter involvert i cellulære responser og PM-reparasjonsmekanismer i levende celler. I tillegg til å studere proteinresponsen ved PM-skade, kan denne tilnærmingen også brukes til lokalt punktering av vesikler, og dermed muliggjøre en undersøkelse av proteinresponsen i både protein-protein og proteinmembrandynamikk. Videre tillater denne metoden en kvantitativ analyse av samspillet mellom proteiner, lipider og små molekyler når membraner forstyrres. Samlet har disse fremskrittene potensial til å kaste lys over noen av de uløste spørsmålene angående det intrikate og komplekse plasmamembranreparasjonsmaskineriet.
