La membrana celular, que sirve como barrera física y plataforma de señalización, es vitalpara la supervivencia celular. A lo largo de todo su ciclo celular, la membrana plasmática (PM) está sometida a daños, como lesiones mecánicas 2,3,4,5 y químicas6 inducidas por estrés. Para mantener la integridad de la membrana y garantizar la supervivencia celular, la célula ha desarrollado mecanismos robustos de reparación de la membrana plasmática (PMR). Estos mecanismos dependen de diversas estrategias, como la reorganización del citoesqueleto, la fusión de membranas y las estrategias de reemplazo de membranas 7,8,9,10,11, todas ellas basadas en el reclutamiento de proteínas específicas. En particular, los miembros de la familia de proteínas de la anexina han sido identificados como proteínas clave asociadas con los procesos de PMR 1,9,12,13,14,15,16. Después de la lesión por PM, la célula experimenta una afluencia de iones de calcio (Ca2+), lo que representa una amenaza inmediata para la supervivencia de la célula17. En respuesta a la afluencia de Ca2+, las proteínas de anexina, que se encuentran predominantemente en el citosol, se unen a la valva interna de la membrana plasmática dañada como parte de las estrategias de PMR18. La anexina A2 (ANXA2) fue uno de los primeros miembros de la familia de las anexinas en asociarse con PMR en la distrofia muscular deficiente en disferlina y se sugirió que mediaba la reparación mediante la fusión de vesículas intracelulares a la PM cerca del sitio de la lesión 5,19,20,21. Posteriormente, se han atribuido varias funciones a las anexinas22, y su papel en la PMR ha atraído una mayor atención en los últimos 20 años. Sin embargo, el papel exacto de las anexinas en la PMR aún no se comprende completamente 15,18,21,22.
Este artículo propone un método para investigar la interacción proteína-membrana y la dinámica de la membrana de una manera controlada y altamente localizada, utilizando una combinación de microscopía confocal, pinzas ópticas y nanopartículas de oro (AuNPs). Este método permite el estudio cuantitativo de las interacciones entre proteínas, lípidos y moléculas pequeñas en respuesta al daño de la membrana y la afluencia de Ca2+ . A pesar de la complejidad y multiplicidad de componentes involucrados en el proceso de reparación de la membrana, se han empleado sistemas de membrana simplificados que imitan la membrana plasmática para obtener una comprensión mecanicista más profunda de la dinámica de la membrana y la respuesta de las proteínas de anexina a la disrupción de la membrana16. Se eligieron vesículas lipídicas unilamelares gigantes (GUV) como sistema modelo de membranas con una composición lipídica especificada. Los GUV se generaron utilizando el método de hidratación asistida por gel, específicamente la hidratación en gel de alcohol polivinílico, según lo descrito por Weinberger et al.23, lo que permitió la encapsulación eficiente de las anexinas en los GUV.
La utilización de la irradiación láser de infrarrojo cercano (NIR) sobre nanopartículas metálicas (NP) induce un calentamiento significativo de la NP, lo que la convierte en un método eficaz para establecer una fuente de calor local explotada en aplicaciones biomédicas24. El método se utilizó inicialmente para medir directamente la temperatura que rodea a un solo AuNP en ensayos biomiméticos 2D y 3D. En estos ensayos25,26, las nanopartículas plasmónicas se irradiaron sobre una bicapa lipídica soportada o se atraparon ópticamente cerca de los GUV que experimentan una transición de fase térmica local tras el calentamiento local, lo que permite cuantificar y controlar el perfil de temperatura exacto alrededor de la partícula. Este perfil de temperatura de referencia se ha utilizado cuando se investigan o manipulan muestras biológicas. Nuevos avances en el método han facilitado la inducción de poros nanoscópicos en las membranas27, lo que permite la fusión de vesículas y células28,29. Otros estudios han investigado el comportamiento de las proteínas de membrana periférica en GUVs29 y proteínas transmembrana30 mediante la creación de nuevas vesículas híbridas, mientras que la administración de fármacos específicos de células también se ha explorado para controlar y estudiar las respuestas celulares o la expresión génica 28,29,31,32,33. Recientemente, el método se ha utilizado para investigar las respuestas de las proteínas al daño de la membrana 32,34,35.
Existen varios métodos para interrumpir la membrana plasmática con el fin de explorar las respuestas celulares y la reparación de la membrana. Estos incluyen punciones con microagujas, agitación de microperlas y raspado celular, todos los cuales pueden alterar mecánicamente la membrana celular 14,36,37. El daño inducido químicamente se puede lograr mediante la adición de detergentes 5,38 o toxinas bacterianas39,40 que desestabilizan la bicapa lipídica y generan poros de membrana a través de la membrana plasmática. Además, las lesiones inducidas ópticamente por láseres de onda continua y pulsados se han utilizado para estudiar los componentes de PMR, como las proteínasde anexina 5,14,21,41, en combinación con nanopartículas plasmónicas 42,43,44,45. A pesar de la eficiencia de los láseres pulsados de alta potencia, pueden causar lesiones y daños significativos en el interior de la célula a lo largo de la trayectoria del haz. Además, los cambios detallados que se producen en la materia biológica tras la irradiación láser pulsada y si crea un poro bien definido aún no se han investigado. En este artículo se presenta un método alternativo que emplea termoplasmónica para inducir agujeros nanoscópicos en el PM de forma controlada34,35 sin causar daños significativos a las estructuras internas. Esto se logra mediante la exposición de NPs plasmónicas a un láser NIR altamente enfocado, lo que resulta en un aumento de temperatura extremadamente localizado que puede alcanzar fácilmente temperaturas superiores a 200 °C, lo que puede conducir a pequeñas explosiones nanoscópicas 25,46,47. Este proceso se puede controlar ajustando la intensidad del láser, así como el tamaño, la forma y la composición de los NP48. Al emplear esta técnica, los investigadores pueden explorar el papel de las proteínas en la reparación de PM en células vivas, lo que podría ayudar a abordar algunas de las preguntas sin respuesta sobre la participación de las proteínas de anexina en la reparación de la membrana sin comprometer la viabilidad celular.
El atrapamiento óptico de nanopartículas plasmónicas ha sido bien establecido por estudios previos 25,49,50,51,52; sin embargo, se pueden obtener conocimientos adicionales sobre las propiedades termoplasmónicas de las nanopartículas 53,54,55 en los materiales suplementarios (Archivo Suplementario 1). El método termoplasmónico se puede utilizar para crear agujeros nanoscópicos en el PM con el fin de estudiar la respuesta celular y los mecanismos de reparación. Más precisamente, la punción se puede lograr mediante el calentamiento óptico de AuNPs en las proximidades de la membrana, como se muestra en las Figuras 1A y B. Esta punción localizada permite la entrada de Ca2+, que fue verificada por un sensor de calcio, activando así la maquinaria PMR. Para los experimentos con células vivas, se inmovilizaron AuNPs con un diámetro de 200 nm en la superficie debajo de la célula para monitorizar el papel de ANXA2 en PMR a través de microscopía confocal. El láser NIR (Figura 1A, B), con una longitud de onda de 1064 nm, irradia el AuNP, explotando sus propiedades plasmónicas (Figura 1C), lo que resulta en un calentamiento local sustancial (Figura 1D) en la ventana de transparencia biológica49 mientras causa un daño mínimo a la propia célula. La región de alta temperatura que rodea el AuNP disminuye rápidamente en un 30-40% a una distancia correspondiente al radio del NP, como se muestra en la Figura 1E, lo que permite una lesión extremadamente confinada en las tres dimensiones.
Expediente Complementario 1. Haga clic aquí para descargar este archivo.
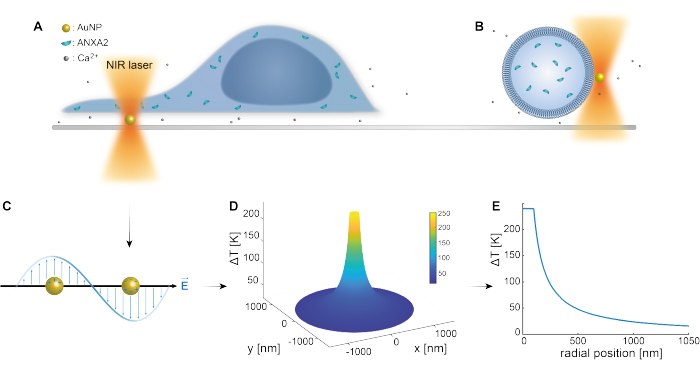
Figura 1: Esquema esquemático del método experimental. (A) las células transfectadas con ANXA están situadas encima de nanopartículas de oro inmovilizadas (AuNPs) en la superficie, o (B) vesículas unilamelares gigantes (GUVs) con ANXAs encapsuladas están suspendidas en un medio que contiene AuNPs. (C) Un solo AuNP es irradiado por la trampa óptica NIR, donde la interacción entre el campo electromagnético entrante y los electrones de conducción conduce a la oscilación colectiva de electrones dentro del NP. (D) Este proceso da como resultado un aumento de temperatura altamente confinado pero significativo. Para estimar la temperatura en la superficie del NP, se emplea la teoría de Mie, y se calcula un perfil de temperatura (E) para un AuNP con un diámetro de 200 nm y una intensidad láser I = 6,36 x 108 W/cm2. Haga clic aquí para ver una versión más grande de esta figura.
Para minimizar el efecto térmico sobre la membrana celular, las AuNP solo se irradian durante ~ 1 segundo. Esto provoca un estallido transitorio y local de calentamiento, lo que reduce el daño a las proteínas que normalmente requieren más tiempo para desplegarse. Tras la punción de la membrana, las proteínas de anexina se reclutan en una fracción de segundo y, en pocos segundos, se forma un andamio en forma de anillo de anexina alrededor del sitio de la lesión (Figura 2). Este enfoque también se ha aplicado para explorar la participación de ANXA5 tanto en células vivas como en membranas modelo16 en un esfuerzo por arrojar luz sobre el esquema completo de los procesos de reparación. Si bien el enfoque principal se ha centrado en el reclutamiento correlacionado de varias proteínas de anexina, los aspectos biofísicos del mecanismo de reparación aún no se han dilucidado.
Para implementar completamente el método propuesto, se requieren tres componentes clave: microscopía confocal, pinzas ópticas y nanopartículas metálicas. Las pinzas ópticas se utilizan para atrapar AuNPs, y su construcción se puede lograr siguiendo el procedimiento descrito por Neuman et al.49. Sin embargo, si la construcción de una pinza óptica resulta ser demasiado difícil, se puede utilizar un láser NIR bien enfocado para irradiar AuNPs inmovilizadas debajo de las células. Si bien se eligieron AuNP esféricas para este protocolo, también se pudo utilizar una variedad de partículas plasmónicas con espectros de absorción sintonizables para lograr un gradiente de temperatura altamente localizado dentro de la región NIR48.
La obtención de imágenes de fluorescencia es necesaria para observar el papel de las proteínas marcadas con fluorescencia y, por lo tanto, la microscopía de reflexión interna total (TIRF)56 podría considerarse como una alternativa a la obtención de imágenes confocal. Sin embargo, esta técnica solo permite obtener imágenes de superficie y no sería compatible con los experimentos de vesículas de membrana modelo. En consecuencia, tanto las pinzas ópticas como el microscopio confocal son esenciales para la localización precisa de la nanopartícula y la investigación detallada del área local que rodea la lesión celular. Para irradiar eficazmente la nanopartícula con un foco láser limitado por difracción, es necesario visualizar la nanopartícula. Esto se puede lograr de manera óptima mediante microscopía de reflexión, que es una característica de imagen estándar de los microscopios confocales Leica. Sin embargo, si no se dispone de imágenes de reflexión o dispersión, se pueden considerar métodos alternativos, como el marcaje fluorescente de AuNP menos eficiente.
En resumen, el método termoplasmónico altamente controlable y localizado presentado en este estudio tiene el potencial de servir como una excelente plataforma para investigar los componentes moleculares involucrados en las respuestas celulares y los mecanismos de reparación de PM en células vivas. Además de estudiar la respuesta de la proteína al daño por PM, este enfoque también se puede utilizar para perforar localmente vesículas, lo que permite una investigación de la respuesta de la proteína tanto en la dinámica proteína-proteína como en la proteína-membrana. Además, este método permite un análisis cuantitativo de las interacciones entre proteínas, lípidos y moléculas pequeñas cuando se rompen las membranas. En conjunto, estos avances tienen el potencial de arrojar luz sobre algunas de las preguntas no resueltas relacionadas con la intrincada y compleja maquinaria de reparación de membranas plasmáticas.
