Cellmembranet, som fungerar både som en fysisk barriär och en signalplattform, är avgörande för cellens överlevnad1. Under hela sin cellcykel utsätts plasmamembranet (PM) för skador, såsom mekaniska 2,3,4,5 och kemiska6 stressinducerade skador. För att upprätthålla membranintegriteten och säkerställa cellens överlevnad har cellen utvecklat robusta mekanismer för reparation av plasmamembran (PMR). Dessa mekanismer beror på olika strategier, såsom omorganisation av cytoskelett, membranfusion och membranersättningsstrategier 7,8,9,10,11, som alla är beroende av rekrytering av specifika proteiner. Noterbart är att medlemmar av annexinproteinfamiljen har identifierats som nyckelproteiner associerade med processerna för PMR 1,9,12,13,14,15,16. Efter partikelskada upplever cellen ett inflöde av kalciumjoner (Ca2+), vilket utgör ett omedelbart hot mot cellens överlevnad17. Som svar på Ca2+-inflöde binder annexinproteiner, som huvudsakligen finns i cytosolen, till det inre bladet i det skadade plasmamembranet som en del av PMR-strategierna18. Annexin A2 (ANXA2) var en av de första medlemmarna i annexinfamiljen som associerades med PMR vid dysferlin-bristfällig muskeldystrofi och föreslogs förmedla reparation genom att smälta intracellulära vesiklar till PM nära skadestället 5,19,20,21. Därefter har flera funktioner tillskrivits annexiner22, och deras roll i PMR har fått ökad uppmärksamhet under de senaste 20 åren. Den exakta rollen för annexiner i PMR är dock fortfarande inte helt klarlagd 15,18,21,22.
Denna artikel föreslår en metod för att undersöka protein-membraninteraktion och membrandynamik på ett kontrollerat och mycket lokaliserat sätt, med hjälp av en kombination av konfokalmikroskopi, optisk pincett och guldnanopartiklar (AuNP). Denna metod möjliggör kvantitativa studier av protein-, lipid- och småmolekylinteraktioner som svar på membranskador och Ca2+ -inflöde. Trots komplexiteten och mångfalden av komponenter som är involverade i processen för membranreparation, har förenklade membransystem som efterliknar plasmamembranet använts för att få en djupare mekanistisk förståelse av membrandynamik och annexinproteiners respons på membranstörningar16. Gigantiska unilamellära lipidvesiklar (GUV) valdes som modellmembransystem med en specificerad lipidsammansättning. GUV:erna genererades med hjälp av den gelassisterade hydratiseringsmetoden, särskilt polyvinylalkoholgelhydratisering, som beskrivs av Weinberger et al.23, vilket möjliggjorde effektiv inkapsling av annexiner i GUV:er.
Användningen av nära-infraröd (NIR) laserbestrålning på metalliska nanopartiklar (NP) inducerar betydande uppvärmning av NP, vilket gör det till en effektiv metod för att etablera en lokal värmekälla som utnyttjas i biomedicinska tillämpningar24. Metoden användes ursprungligen för att direkt mäta temperaturen kring en enda AuNP i både 2D och 3D biomimetiska analyser. I dessa analyser25,26 bestrålades de plasmoniska nanopartiklarna på ett lipiddubbelskikt eller optiskt fångade nära GUV:er som genomgick en lokal termisk fasövergång vid lokal uppvärmning, vilket möjliggjorde kvantifiering och kontroll av den exakta temperaturprofilen runt partikeln. Denna referenstemperaturprofil har använts vid undersökning eller manipulering av biologiska prover. Ytterligare framsteg i metoden har underlättat induktionen av nanoskopiska porer i membran27, vilket möjliggör vesikel- och cellfusion28,29. Andra studier har undersökt beteendet hos perifera membranproteiner i GUVs29 och transmembranproteiner30 genom att skapa nya hybridvesiklar, medan cellspecifik läkemedelsleverans också har utforskats för att kontrollera och studera cellulära svar eller genuttryck 28,29,31,32,33. På senare tid har metoden använts för att undersöka proteiners respons på membranskador 32,34,35.
Det finns flera metoder för att störa plasmamembranet för att utforska cellulära svar och membranreparation. Dessa inkluderar mikronålspunkteringar, skakning av mikropärlor och cellskrapning, som alla kan störa cellmembranet mekaniskt 14,36,37. Kemiskt inducerad skada kan uppnås genom tillsats av rengöringsmedel 5,38 eller bakterietoxiner39,40 som destabiliserar lipiddubbelskiktet och genererar membranporer över plasmamembranet. Dessutom har optiskt inducerade skador av kontinuerliga vågor och pulsade lasrar använts för att studera PMR-komponenter, såsom annexinproteiner 5,14,21,41, i kombination med plasmoniska nanopartiklar 42,43,44,45. Trots effektiviteten hos pulsade lasrar med hög effekt kan de orsaka betydande skador och skador på cellens inre längs strålvägen. Dessutom återstår det att undersöka de detaljerade förändringar som sker i det biologiska materialet vid pulsad laserbestrålning och om det skapar en väldefinierad por. En alternativ metod presenteras i denna artikel, som använder termoplasmonik för att inducera nanoskopiska hål i PM på ett kontrollerat sätt34,35 utan att orsaka betydande skada på de inre strukturerna. Detta åstadkoms genom att exponera plasmoniska NP för en mycket fokuserad NIR-laser, vilket resulterar i en extremt lokal temperaturökning som lätt kan nå temperaturer över 200 °C, vilket kan leda till små nanoskopiska explosioner 25,46,47. Denna process kan styras genom att justera laserintensiteten samt storleken, formen och sammansättningen av NP48. Genom att använda denna teknik kan forskare utforska proteinernas roll i PM-reparation i levande celler, vilket kan hjälpa till att ta itu med några av de obesvarade frågorna om annexinproteiners inblandning i membranreparation utan att äventyra cellviabiliteten.
Den optiska infångningen av plasmoniska nanopartiklar är väl etablerad i tidigare studier 25,49,50,51,52; Ytterligare insikter om de termoplasmoniska egenskaperna hos nanopartiklarna 53,54,55 kan dock erhållas i de kompletterande materialen (Supplementary File 1). Den termoplasmoniska metoden kan användas för att skapa nanoskopiska hål i PM i syfte att studera cellulära respons- och reparationsmekanismer. Mer exakt kan punkteringen åstadkommas genom optisk uppvärmning av AuNP i närheten av membranet, som visas i figurerna 1A och B. Denna lokaliserade punktering möjliggör Ca2+-inflöde, vilket verifierades av en kalciumsensor, vilket aktiverar PMR-maskineriet. För experiment med levande celler immobiliserades AuNP med en diameter på 200 nm på ytan under cellen för att övervaka ANXA2:s roll i PMR via konfokalmikroskopi. NIR-lasern (figur 1A,B), med en våglängd på 1064 nm, bestrålar AuNP och utnyttjar dess plasmoniska egenskaper (figur 1C), vilket resulterar i betydande lokal uppvärmning (figur 1D) i det biologiska transparensfönstret49 samtidigt som den orsakar minimal skada på själva cellen. Högtemperaturområdet som omger AuNP minskar snabbt med 30-40 % på ett avstånd som motsvarar NP:s radie, som visas i figur 1E, vilket möjliggör en extremt begränsad skada i alla tre dimensionerna.
Kompletterande fil 1. Klicka här för att ladda ner den här filen.
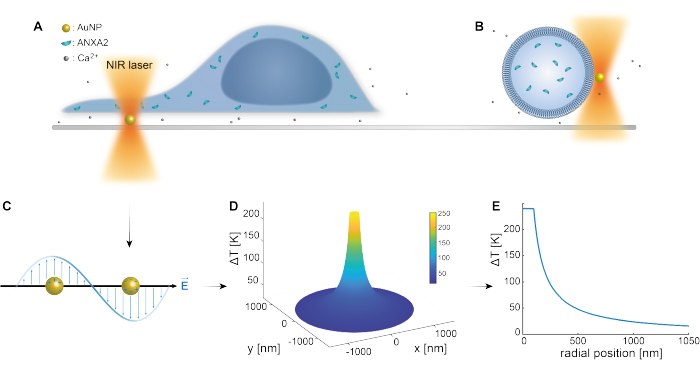
Figur 1: Schematisk översikt över den experimentella metoden. (A) ANXA-transfekterade celler är belägna ovanpå immobiliserade guldnanopartiklar (AuNP) på ytan, eller (B) gigantiska unilamellära vesiklar (GUV) med inkapslad ANXA är suspenderade i ett medium som innehåller AuNPs. (C) En enda AuNP bestrålas av den optiska NIR-fällan, där interaktionen mellan det inkommande elektromagnetiska fältet och ledningselektronerna leder till en kollektiv svängning av elektroner inom NP. (D) Denna process resulterar i en mycket begränsad men ändå betydande temperaturökning. För att uppskatta temperaturen vid NP-ytan används Mie-teorin, och en (E) temperaturprofil beräknas för en AuNP med en diameter på 200 nm och laserintensiteten I = 6,36 x 108 W/cm2. Klicka här för att se en större version av denna figur.
För att minimera den termiska effekten på cellmembranet bestrålas AuNP endast i ~1 sekund. Detta orsakar en övergående och lokal explosion av uppvärmning, vilket minskar skadorna på proteiner som vanligtvis kräver mer tid för att utvecklas. Vid membranpunktion rekryteras annexinproteiner på bråkdelen av en sekund, och inom några sekunder bildas en annexinringliknande ställning runt skadestället (Figur 2). Detta tillvägagångssätt har också tillämpats för att utforska involveringen av ANXA5 i både levande celler och modellmembran16 i ett försök att belysa hela schemat för reparationsprocesserna. Medan det primära fokuset har legat på korrelerande rekrytering av olika annexinproteiner, har de biofysiska aspekterna av reparationsmekanismen ännu inte klarlagts.
För att fullt ut implementera den föreslagna metoden krävs tre nyckelkomponenter: konfokalmikroskopi, optisk pincett och metallnanopartiklar. Optisk pincett används för att fånga AuNPs, och deras konstruktion kan uppnås genom att följa proceduren som beskrivs av Neuman et al.49. Men om det visar sig vara för utmanande att bygga en optisk pincett, kan en tätt fokuserad NIR-laser användas för att bestråla AuNPs immobiliserade under cellerna. Medan sfäriska AuNP valdes för detta protokoll, kunde en mängd plasmoniska partiklar med avstämbara absorptionsspektra också användas för att uppnå en mycket lokal temperaturgradient inom NIR-regionen48.
Fluorescensavbildning är nödvändig för att observera de fluorescerande märkta proteinernas roll, och därför kan total intern reflektionsmikroskopi (TIRF)56 övervägas som ett alternativ till konfokal avbildning. Denna teknik tillåter dock endast ytavbildning och skulle inte vara kompatibel med modellmembranvesikelexperimenten. Följaktligen är både den optiska pincetten och konfokalmikroskopet viktiga för den exakta lokaliseringen av nanopartikeln och detaljerad undersökning av det lokala området kring cellskadan. För att effektivt bestråla nanopartikeln med ett diffraktionsbegränsat laserfokus är det nödvändigt att visualisera nanopartikeln. Detta kan uppnås optimalt genom reflektionsmikroskopi, som är en standardavbildningsfunktion för Leicas konfokalmikroskop. Om reflektions- eller spridningsavbildning inte är tillgänglig kan dock alternativa metoder, såsom den mindre effektiva fluorescerande AuNP-märkningen, övervägas.
Sammanfattningsvis har den mycket kontrollerbara och lokaliserade termoplasmoniska metoden som presenteras i denna studie potential att fungera som en utmärkt plattform för att undersöka de molekylära komponenter som är involverade i cellulära svar och PM-reparationsmekanismer i levande celler. Förutom att studera proteinresponsen vid PM-skada kan detta tillvägagångssätt också användas för att lokalt punktera vesiklar, vilket möjliggör en undersökning av proteinresponsen i både protein-protein och protein-membrandynamik. Dessutom möjliggör denna metod en kvantitativ analys av interaktionerna mellan proteiner, lipider och små molekyler när membran störs. Sammantaget har dessa framsteg potential att kasta ljus över några av de olösta frågorna om det intrikata och komplexa maskineriet för reparation av plasmamembran.
