This protocol evaluates the capacity of sEVs derived from placentas cultured in hypoxia to disrupt the BBB in nonpregnant mice. This method allows one to understand better the potential connection between the placenta and the brain in normal and pathological conditions. In particular, this method may constitute a proxy to analyze placental sEVs participation in the onset of cerebral complications in preeclampsia.
Contrary to mice injected with sEVs-Nor, mice injected with sEVs-Hyp show a progressive decline in the neurological score until 24 h (Table 1), which suggests the sEVs-Hyp capacity to impair brain function.
Also, mice brains of the sEVs-Hyp injected group have higher fresh weight than those isolated from mice injected with sEVs-Nor or control mice (0.51 ± 0.008; 0.46 ± 0.008; 0.47 ± 0.01 g, respectively), which may constitute a gross indicator of brain edema45.
Compatible with this finding, this protocol allows one to identify Evan's blue extravasation as an indicator of disruption of the BBB. In that regard, brains from mice injected with sEVs-Hyp have higher Evan's blue extravasation than brains from the sEVs-Nor group (Figure 5A).
Although the underlying mechanism of disruption of the BBB induced by sEVs-Hyp was not analyzed with this protocol, results also indicate that sEVs-Hyp injected mice showed reduced protein amounts of CLND-5 in the areas in which the BBB was most affected (i.e., posterior areas) (Figure 5B). Therefore, it is feasible that sEVs-hyp impairs the expression of the function of this critical endothelial tight junction protein.
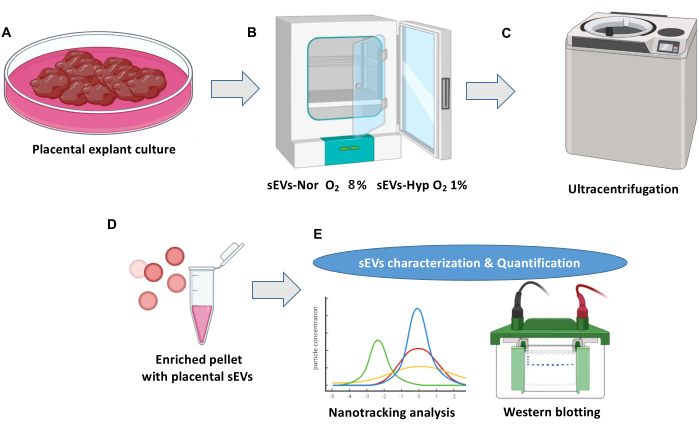
Figure 1: Placental explant culture and extracellular vesicle isolation protocol. (A) Normal placenta explant cultures. (B) Explants are distributed into two conditions for the biogenesis of placental small extracellular vesicles (sEVs). Normoxia (sEVs-Nor, 8% O2) or hypoxia (sEVs-Hyp, 1% O2) for 18 h. (C) Conditioned media is harvested, filtered, and centrifuged to eliminate cell debris. (D) sEVs are isolated by ultra-centrifugations. (E) sEVs are characterized using nano-tracking analysis and western blot. Please click here to view a larger version of this figure.
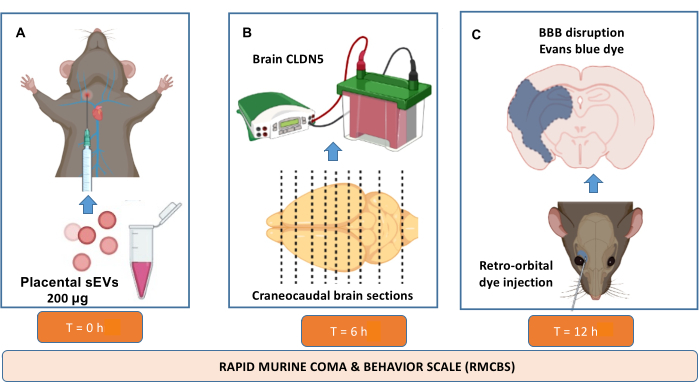
Figure 2: In vivo evaluation of blood-brain barrier disruption protocol. Nonpregnant C57BL6/J mice, 4-6 months old, are used. (A) Via external jugular vein, animals received sEVs (200 μg of total protein) isolated from Normoxic (sEVs-Nor, 8% O2) or hypoxic (sEVs-Hyp, 1% O2) placental cultures. RMCBS is monitored at 0-24 h after injection. (B) 6 h after sEVs injection, brains are extracted and sectioned into nine segments for protein extraction. Claudin 5 (CLDN5) is analyzed in homogenates of those nine sections. (C) Evan's blue extravasation analysis (24 h after sEVs injection) was analyzed after retro-orbital puncture injection in each of the nine segments. Please click here to view a larger version of this figure.
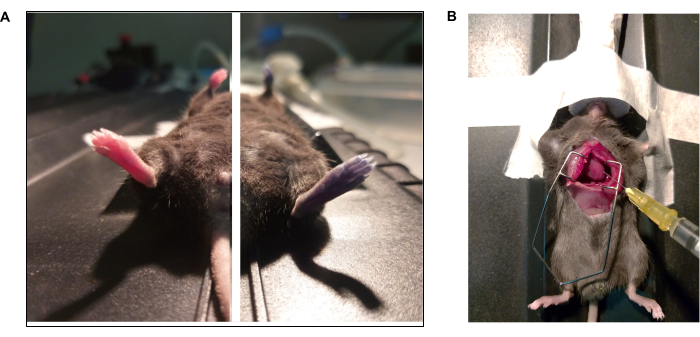
Figure 3: Photo documentaries of Evan' s blue dye and intracardial perfusion protocol. (A) Mouse received Evan's blue via retro-orbital injection. (Left) Animal before and (Right) after injection (15 s) of Evan's blue. (B) Thoracotomy to perform intracardial perfusion of phosphate buffer solution (1x PBS) and paraformaldehyde (4% PFA). The left ventricle is pointed with the tip of the needle. Please click here to view a larger version of this figure.
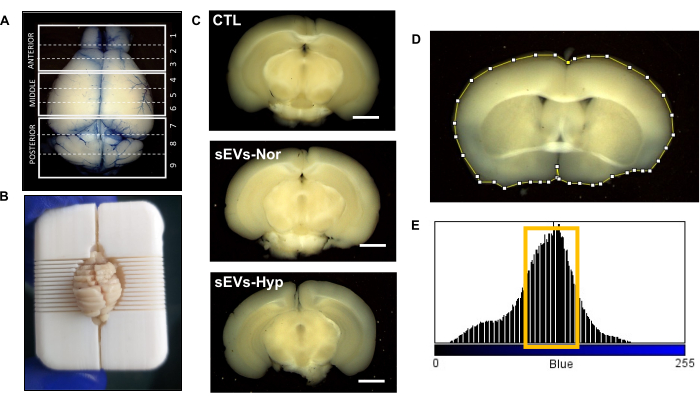
Figure 4: Analysis of Evan's blue extravasation after injection of sEVs. (A) Representative image of the whole brain showing Evan's blue extravasation. The dashed line represents nine sections obtained from the whole brain. (B) Brain dissected using a brain mice slicer. (C) Representative images of brain slices at 24 h after sEVs injection. Control (CTL), placenta in normoxic (sEVs-Nor) or hypoxic conditions (sEVs-Hyp). Scale bar = 0.4 cm. (D) Digital outlining of brain slice using ImageJ. (E) Histogram in the blue channel. Values between 75-110 are associated with Evan's blue extravasation. Please click here to view a larger version of this figure.
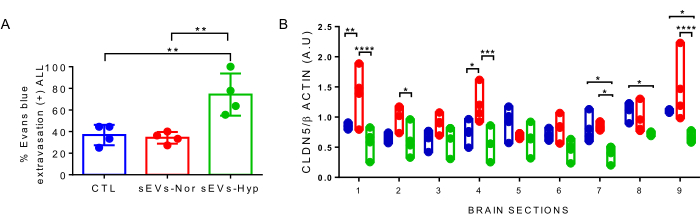
Figure 5: Evan's blue extravasation and claudin-5 levels in mice injected with sEVs isolated from placental explants. (A) Percentage of Evan's blue (EB) extravasation considering the whole brain sections. Control (CTL, blue), placenta in normoxic (sEVs-Nor, red) or hypoxic conditions (sEVs-Hyp, green). (B) Relative levels of claudin 5 (CLDN5) in the nine brain sections were obtained from the three experimental groups. β-actin is used as a loading control. Values are mean ± interquartile range. Each dot represents an individual experimental subject. *p < 0.05, **p < 0.005. ****p < 0.0001. ***p < 0.001; ANOVA test followed by Bonferroni post-test. Please click here to view a larger version of this figure.
| Time (h) | Control | sEVs-Normoxia | sEVs-Hypoxia | ANOVA |
| 0 | 18.75 ± 0.250 | 18.5 ± 0.288 | 18.75 ± 0.250 | ns |
| 3 | 18.5 ± 0.866 | 17 ± 0.707 | 13.25 ± 1.750*α | 0.006 |
| 6 | 19.25 ± 0.750 | 17 ± 0.577 | 11.75 ± 1.250*α | 0.002 |
| 12 | 18.5 ± 0.645 | 16.75 ± 1.109 | 13 ± 0.816*α | 0.001 |
| 24 | 19.5 ± 0.288 | 17.75 ± 0.250 | 10.25 ± 0.853*α | <0.0001 |
| *p < 0.01 versus control. αp < 0.01 versus sEVs-Normoxia | ||||
Table 1: Rapid murine coma and behavior scale (RMCBS) after 24 h post sEVs injection. The score is expressed as mean ± SEM. Animals' scores closest to 20 are standard, while the lower the score, the higher the dysfunction of the CNS.
