תחום הביולוגיה הסינתטית מלמטה למעלה מתמקד בבניית תאים (מינימליים) 1,2 וביוריאקטורים מטבוליים למטרות ביוטכנולוגיות 3,4 או ביו-רפואיות 5,6,7,8. בניית תאים סינתטיים מספקת פלטפורמה ייחודית המאפשרת לחוקרים לחקור חלבונים (ממברנה) בתנאים מוגדרים היטב המחקים את אלה של סביבות טבעיות, ומאפשרת גילוי תכונות מתפתחות ופונקציות ביוכימיות נסתרות של חלבונים ורשתות תגובה9. כצעד ביניים לקראת תא סינתטי המתפקד באופן אוטונומי, מפותחים מודולים הלוכדים תכונות חיוניות של תאים חיים כגון שימור אנרגיה מטבולית, סינתזת חלבונים ושומנים, והומאוסטזיס. מודולים כאלה לא רק לשפר את ההבנה שלנו של החיים, אלא גם יש יישומים פוטנציאליים בתחומי רפואה8 וביוטכנולוגיה10.
חלבונים טרנסממברנליים נמצאים בלב כמעט כל רשת מטבולית מכיוון שהם מעבירים מולקולות לתוך התא או החוצה ממנו, מאותתים ומגיבים לאיכות הסביבה, וממלאים תפקידים ביוסינתטיים רבים. לפיכך, הנדסה של מודולים מטבוליים בתאים סינתטיים דורשת ברוב המקרים הרכבה מחדש של חלבוני ממברנה אינטגרליים ו / או היקפיים לתוך דו-שכבה קרום המורכבת מליפידים ספציפיים ושלמות גבוהה (חדירות נמוכה). הטיפול בחלבוני ממברנה אלה מאתגר ודורש ידע ספציפי ומיומנויות ניסוייות.
מספר שיטות פותחו כדי לשחזר חלבוני ממברנה בתוך שלפוחיות פוספוליפידים, לרוב במטרה לחקור את הפונקציה11,12, תקנה13, תכונות קינטיות14,15, תלות שומנים15,16, ו / או יציבות17 של חלבון מסוים. שיטות אלה כוללות דילול מהיר של חלבון המסיס בדטרגנטים למצע מימי בנוכחות ליפידים18, סילוק דטרגנטים על ידי דגירה של חלבון מסיס בדטרגנטים עם בועיות שומנים מעורערות דטרגנטים וספיגת הדטרגנטים על חרוזי פוליסטירן19, או הסרת דטרגנטים על ידי דיאליזה או כרומטוגרפיה של אי הכללת גודל20. ממיסים אורגניים שימשו ליצירת בועיות שומנים, למשל, באמצעות היווצרות שלבי שמן-מים21, אך רוב חלבוני הממברנה האינטגרלית מושבתים כאשר הם נחשפים לממיסים כאלה.
במעבדה שלנו, אנו בעיקר משחזרים חלבוני ממברנה בשיטת ספיגת חומרי ניקוי ליצירת שלפוחיות חד-למלריות גדולות (LUVs)19. שיטה זו מאפשרת הרכבה מחדש של חלבוני ממברנה מרובים ואת האנקפסולציה בלומן השלפוחית של אנזימים, מטבוליטים ובדיקות22,23. ניתן להמיר את ה-LUV המכילים חלבון ממברנה לשלפוחיות ענקיות-חד-לאומיות (GUVs) עם/בלי אנקפסולציה של רכיבים מסיסים במים, באמצעות אלקטרופורמציה24 או נפיחות25 בסיוע ג’ל ותנאים ספציפיים לשמירה על שלמות חלבוני הממברנה26.
מאמר זה מציג פרוטוקול להרכבה מחדש ב-LUV של רשת מטבולית מחוץ לשיווי משקל המחדשת ATP באמצעות פירוק L-ארגינין ל-L-אורניתין27. היווצרות ATP קשורה לייצור גליצרול-3-פוספט (G3P), אבן בניין חשובה לסינתזה של פוספוליפידים22,28. המסלול המטבולי מורכב משני חלבוני ממברנה אינטגרליים, ארגינין/אורניתין (ArcD) ואנטיפורטר G3P/Pi (GlpT). נוסף על כך, שלושה אנזימים מסיסים (ArcA, ArcB, ArcC) נדרשים למחזור ATP, ו-GlpK משמש להמרת גליצרול לגליצרול 3-פוספט, באמצעות ATP מפירוק L-ארגינין, ראו איור 1 לסקירה סכמטית של המסלול. פרוטוקול זה מייצג נקודת התחלה טובה לבנייה עתידית של רשתות תגובה מורכבות עוד יותר – לסינתזה של שומנים או חלבונים או לחלוקת תאים. הרכב השומנים של השלפוחיות תומך בפעילות של מגוון רחב של חלבוני ממברנה אינטגרליים והותאם להובלת מולקולות מגוונות לתוך או מחוץ לשלפוחיות 27,29,30.
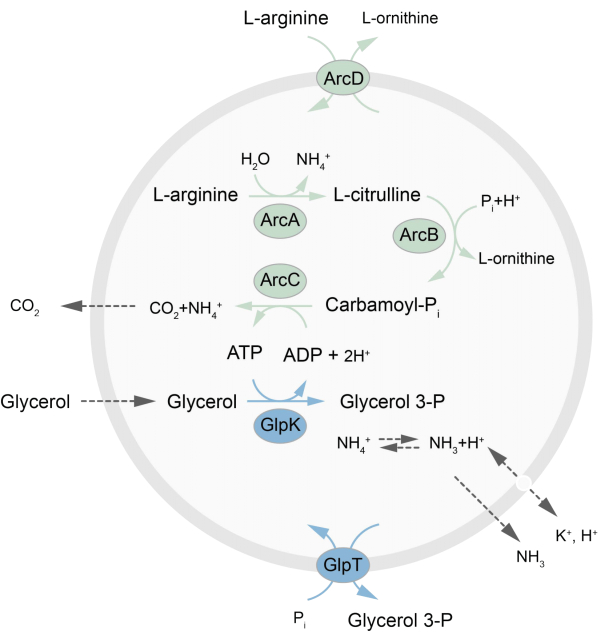
איור 1: סקירה כללית של המסלול לייצור ATP ולסינתזה והפרשה של גליצרול 3-פוספט. לחץ כאן כדי להציג גרסה גדולה יותר של איור זה.
בקיצור, חלבוני ממברנה מטוהרים (מסיסים בדודציל-β-D-מלטוזיד, DDM) מתווספים לבועיות שומנים מוכנות מראש שהתערערו עם טריטון X-100, המאפשר החדרת החלבונים לממברנה. מולקולות הדטרגנט מוסרות לאחר מכן (לאט) על ידי תוספת של חרוזי פוליסטירן פעילים, וכתוצאה מכך נוצרות פרוטאוליפוזומים אטומים היטב. לאחר מכן ניתן להוסיף רכיבים מסיסים לשלפוחיות ולעטוף אותם באמצעות מחזורי הקפאה-הפשרה, אשר לוכדים את המולקולות בתהליך של איחוי ממברנה. השלפוחיות המתקבלות הטרוגניות מאוד ורבות מהן מולטילמלריות. לאחר מכן הם מושחלים דרך מסנן פוליקרבונט עם גודל נקבוביות של 400, 200 או 100 ננומטר, אשר מניב שלפוחיות בגודל אחיד יותר; ככל שהנקבוביות קטנות יותר, כך השלפוחיות הומוגניות וחד-לאומיות יותר, אך במחיר של נפח פנימי קטן יותר. חלבונים שאינם מאוגדים ומולקולות קטנות מוסרים מהתמיסה החיצונית על ידי כרומטוגרפיה של אי הכללת גודל. ניתן להמיר את ה-proteoLUVs לבועיות בגודל מיקרומטר על ידי נפיחות בעזרת ג’ל, ו-proteoGUV אלה נאספים ונלכדים בשבב מיקרופלואידי לצורך אפיון מיקרוסקופי ומניפולציה. איור 2 מציג סקירה סכמטית של הפרוטוקול המלא.
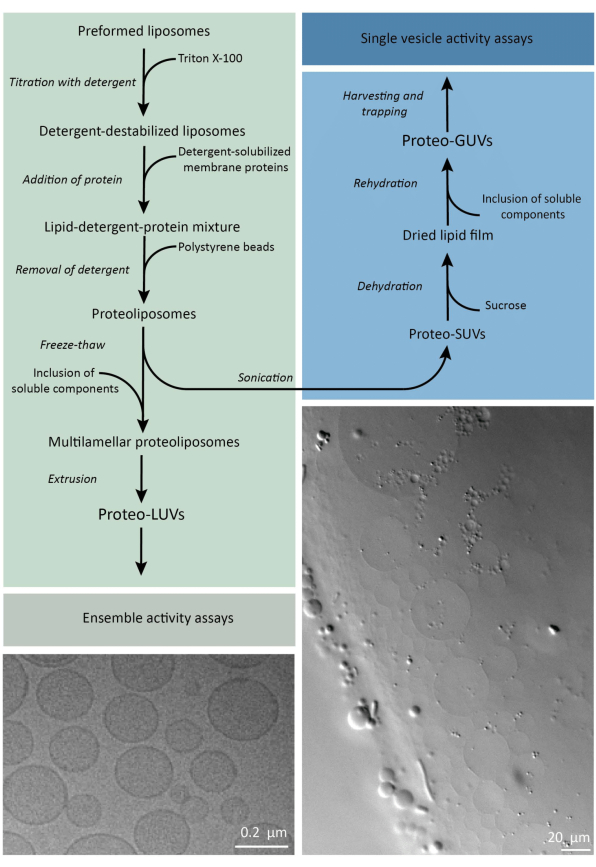
איור 2: סקירה כללית של פרוטוקול להרכבה מחדש של חלבוני ממברנה ועטיפת אנזימים ורכיבים מסיסים במים בשלפוחיות שומנים בגודל תת-מיקרומטר (LUV) ומיקרומטר (GUV). לחץ כאן כדי להציג גרסה גדולה יותר של איור זה.
פרוטוקולי ההרכבה מחדש והאנקפסולציה עובדים היטב והפונקציונליות של החלבונים נשמרת, אך הפרוטיאו-לובים והפרוטאוגו-גובים הם הטרוגניים בגודלם. גישות מיקרופלואידיות31,32 מאפשרות היווצרות שלפוחיות בגודל מיקרומטרי שהן הומוגניות יותר בגודלן, אך הרכבה מחדש פונקציונלית של חלבוני ממברנה אינה אפשרית בדרך כלל מכיוון שממס שיורי בדו-שכבה משבית את החלבונים. גודלם של הפרוטאולובים נע בין 100 ל-400 ננומטר, ובריכוזים נמוכים של אנזימים, האנקפסולציה עלולה להוביל לבועיות עם מסלולים מטבוליים לא שלמים (השפעות סטוכסטיות; ראו איור 3). LUVs אידיאליים לבניית מודולים מטבוליים ספציפיים, כפי שמוצג כאן לייצור ATP ואבני בניין כמו G3P. proteoLUVs כאלה יכולים להיות עטופים באופן פוטנציאלי ב- GUV ולשמש כתאים דמויי אברונים עבור שלפוחיות המאכסן.
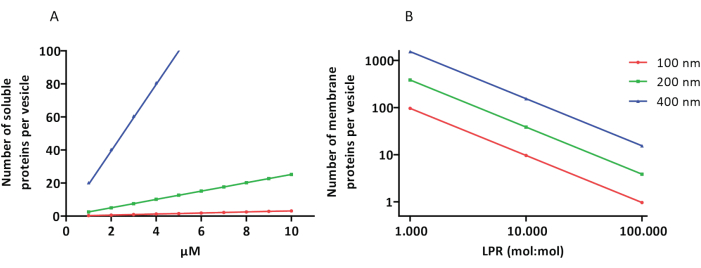
איור 3: מספר המולקולות לשלפוחית בקוטר של 100, 200 או 400 ננומטר. (A) כאשר החלבונים העטוף (אנזימים, גשושיות) נמצאים בטווח של 1-10 מיקרומטר. (B) הבנייה מחדש נעשית ב-1 עד 1,000, 1 עד 10,000, ו-1 עד 100,000 חלבוני ממברנה לכל ליפיד (mol/mol). אנו יוצאים מנקודת הנחה שהמולקולות עטופות בריכוזים שצוינו ומשולבות בממברנה ביחסי חלבון-שומנים אלה. עבור אנזימים מסוימים, ראינו כי הם נקשרים לממברנות, אשר יכול להגדיל את הריכוז לכאורה שלהם שלפוחיות. קיצור: LPR = יחס שומנים-חלבונים אנא לחץ כאן כדי להציג גרסה גדולה יותר של איור זה.
