Aşağıdan yukarıya sentetik biyoloji alanı, biyoteknolojik 3,4 veya biyomedikal amaçlar 5,6,7,8 için (minimal) hücreler 1,2 ve metabolik biyoreaktörler oluşturmaya odaklanır. Sentetik hücrelerin inşası, araştırmacıların, doğal ortamların proteinlerini taklit eden iyi tanımlanmış koşullarda proteinleri (zar) incelemelerine olanak tanıyan, proteinlerin ve reaksiyon ağlarının ortaya çıkan özelliklerinin ve gizli biyokimyasal işlevlerinin keşfedilmesini sağlayan benzersiz bir platform sağlar9. Otonom olarak işleyen bir sentetik hücreye doğru bir ara adım olarak, canlı hücrelerin metabolik enerji korunumu, protein ve lipid sentezi ve homeostaz gibi temel özelliklerini yakalayan modüller geliştirilmiştir. Bu tür modüller sadece yaşam anlayışımızı geliştirmekle kalmaz, aynı zamanda tıp8 ve biyoteknoloji10 alanlarında da potansiyel uygulamalara sahiptir.
Transmembran proteinler, molekülleri hücrenin içine veya dışına taşıdıkları, sinyal verdikleri ve çevrenin kalitesine yanıt verdikleri ve çok sayıda biyosentetik rol oynadıkları için hemen hemen her metabolik ağın merkezinde yer alır. Bu nedenle, sentetik hücrelerdeki metabolik modüllerin mühendisliği, çoğu durumda integral ve/veya periferik membran proteinlerinin, spesifik lipitlerden ve yüksek bütünlükten (düşük geçirgenlik) oluşan bir membran çift tabakasına yeniden yapılandırılmasını gerektirir. Bu zar proteinlerinin kullanımı zordur ve özel bilgi ve deneysel beceriler gerektirir.
Fosfolipid veziküller içindeki zar proteinlerini yeniden oluşturmak için, çoğunlukla spesifik bir proteinin fonksiyon11,12, regülasyon13, kinetik özellikler14,15, lipid bağımlılığı15,16 ve/veya stabilite17’yi incelemek amacıyla çeşitli yöntemler geliştirilmiştir. Bu yöntemler, deterjanla çözündürülmüş proteininlipitlerin 18 varlığında sulu ortama hızlı bir şekilde seyreltilmesini, deterjanla çözündürülmüş proteinin deterjanla destabilize edilmiş lipid vezikülleri ile inkübe edilmesiyle deterjanların uzaklaştırılmasını ve deterjan(lar)ın polistiren boncuklar19 üzerine emilmesini veya deterjanların diyaliz veya boyut dışlama kromatografisi20 ile uzaklaştırılmasını içerir. Organik çözücüler, örneğin yağ-su interfazlarının21 oluşumu yoluyla lipid vezikülleri oluşturmak için kullanılmıştır, ancak integral membran proteinlerinin çoğu, bu tür çözücülere maruz kaldıklarında inaktive edilir.
Laboratuvarımızda, büyük unilameller veziküller (LUV’ler) oluşturmak için çoğunlukla membran proteinlerini deterjan emme yöntemiyle yeniden sulandırıyoruz19. Bu yöntem, çoklu zar proteinlerinin birlikte sulandırılmasına ve enzimlerin, metabolitlerin ve probların vezikül lümeninde kapsüllenmesine izin verir22,23. Membran proteini içeren LUV’ler, elektroformasyon24 veya jel destekli şişme25 ve membran proteinlerinin26 bütünlüğünü korumak için özel koşullar kullanılarak, suda çözünür bileşenlerin kapsüllenmesi ile / kapsüllenmemesi ile dev unilamellar veziküllere (GUV’ler) dönüştürülebilir.
Bu makale, L-arginin’in L-ornitin27’ye parçalanması yoluyla ATP’yi yeniden üreten denge dışı bir metabolik ağın LUV’lerinde sulandırılması için bir protokol sunmaktadır. ATP oluşumu, fosfolipid sentezi için önemli bir yapı taşı olan gliserol-3-fosfat (G3P) üretimi ile birleştirilir22,28. Metabolik yol, bir arginin/ornitin (ArcD) ve bir G3P/Pi antiporter (GlpT) olmak üzere iki integral membran proteininden oluşur. Ek olarak, ATP’nin geri dönüşümü için üç çözünür enzim (ArcA, ArcB, ArcC) gereklidir ve GlpK, L-arginin’in parçalanmasından elde edilen ATP’yi kullanarak gliserolü gliserol 3-fosfata dönüştürmek için kullanılır, yolun şematik bir genel bakışı için Şekil 1’e bakın. Bu protokol, lipitlerin veya proteinlerin sentezi veya hücrelerin bölünmesi için daha da karmaşık reaksiyon ağlarının gelecekteki inşası için iyi bir başlangıç noktasını temsil eder. Veziküllerin lipid bileşimi, çok çeşitli integral membran proteinlerinin aktivitesini destekler ve çeşitli moleküllerin veziküllerin içine veya dışına taşınması için optimize edilmiştir 27,29,30.
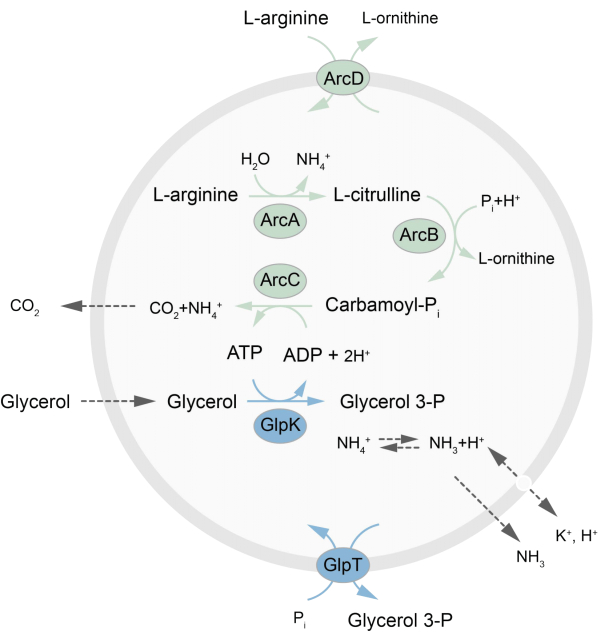
Şekil 1: ATP üretimi ve gliserol 3-fosfat sentezi ve atılımı için yola genel bakış. Bu rakamın daha büyük bir sürümünü görüntülemek için lütfen buraya tıklayın.
Kısacası, saflaştırılmış zar proteinleri (dodesil-β-D-maltosit, DDM içinde çözündürülmüş), proteinlerin zara yerleştirilmesine izin veren Triton X-100 ile destabilize edilmiş önceden oluşturulmuş lipid veziküllerine eklenir. Deterjan molekülleri daha sonra (yavaşça) aktive edilmiş polistiren boncukların eklenmesiyle uzaklaştırılır ve bu da iyi kapatılmış proteolipozomların oluşumuna neden olur. Çözünür bileşenler daha sonra veziküllere eklenebilir ve molekülleri membran füzyonu sürecinde yakalayan donma-çözülme döngüleri yoluyla kapsüllenebilir. Elde edilen veziküller oldukça heterojendir ve çoğu çok lamelli boyalıdır. Daha sonra, daha düzgün boyutta veziküller veren 400, 200 veya 100 nm gözenek boyutuna sahip bir polikarbonat filtreden ekstrüde edilirler; Gözenek boyutu ne kadar küçükse, veziküller o kadar homojen ve tek tiptir, ancak daha küçük bir iç hacim pahasına. Dahil edilmemiş proteinler ve küçük moleküller, boyut dışlama kromatografisi ile harici çözeltiden uzaklaştırılır. ProteoLUV’lar, jel destekli şişme ile mikrometre boyutunda veziküllere dönüştürülebilir ve bu proteoGUV’lar daha sonra toplanır ve mikroskobik karakterizasyon ve manipülasyon için bir mikroakışkan çip içinde tutulur. Şekil 2 , tam protokolün şematik bir genel bakışını göstermektedir.
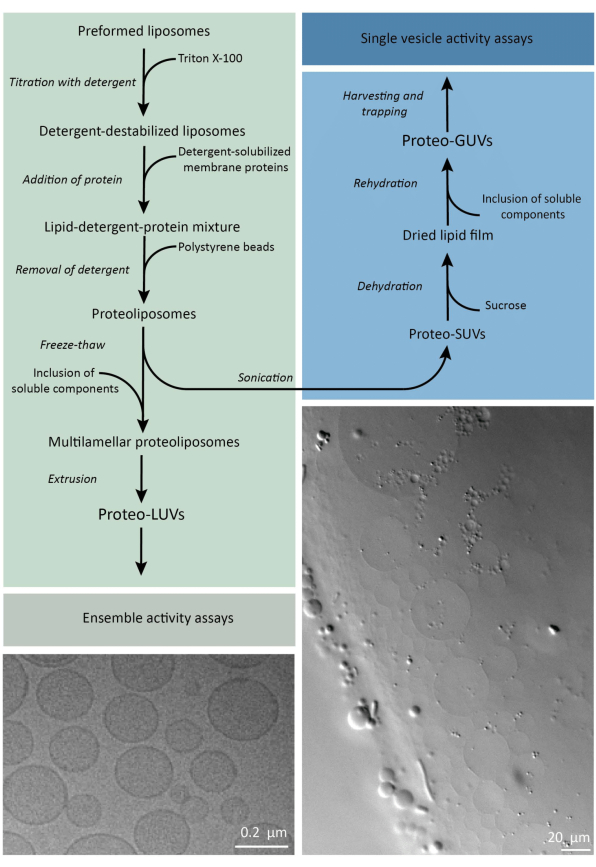
Şekil 2: Mikrometre altı (LUV’ler) ve mikrometre boyutundaki (GUV’ler) lipid veziküllerinde membran proteinlerinin yeniden yapılandırılması ve enzimlerin ve suda çözünür bileşenlerin kapsüllenmesi için protokole genel bakış. Bu rakamın daha büyük bir sürümünü görüntülemek için lütfen buraya tıklayın.
Sulandırma ve kapsülleme protokolleri iyi çalışır ve proteinlerin işlevselliği korunur, ancak proteoLUV’lar ve proteoGUV’lar boyut olarak heterojendir. Mikroakışkan yaklaşımlar 31,32, boyut olarak daha homojen olan mikrometre boyutunda veziküllerin oluşumuna izin verir, ancak zar proteinlerinin işlevsel olarak yeniden yapılandırılması genellikle mümkün değildir, çünkü çift tabakadaki artık çözücü proteinleri inaktive eder. ProteoLUV’lerin boyutları 100 ila 400 nm arasında değişir ve düşük enzim konsantrasyonlarında, kapsülleme eksik metabolik yollara sahip veziküllere yol açabilir (stokastik etkiler; bkz. Şekil 3). LUV’ler, ATP ve G3P gibi yapı taşlarının üretimi için burada gösterildiği gibi belirli metabolik modüller oluşturmak için idealdir. Bu tür proteoLUV’lar potansiyel olarak GUV’lerde kapsüllenebilir ve konakçı veziküller için organel benzeri bölmeler olarak hizmet edebilir.
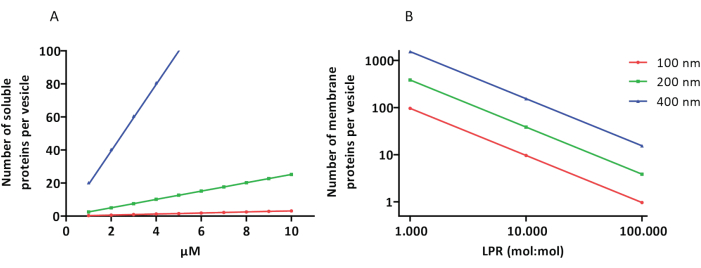
Şekil 3: Çapı 100, 200 veya 400 nm olan vezikül başına molekül sayısı. (A) Kapsüllenmiş proteinler (enzimler, problar) 1-10 μM aralığında olduğunda. (B) Sulandırma, lipid başına 1 ila 1.000, 1 ila 10.000 ve 1 ila 100.000 membran proteini (mol / mol) arasında yapılır. Moleküllerin belirtilen konsantrasyonlarda kapsüllendiği ve bu protein-lipit oranlarında zara dahil edildiği varsayımını yapıyoruz. Bazı enzimler için, zarlara bağlandıklarını gördük, bu da veziküllerdeki görünür konsantrasyonlarını artırabilir. Kısaltma: LPR = Lipid-Protein-Oranı Bu rakamın daha büyük bir sürümünü görüntülemek için lütfen buraya tıklayın.
