Isolation and identification of fungal isolates from M. alternatus
With the aid of attractant traps, a large number (approximately 500 beetles in total) of M. alternatus were collected from five geographical regions. Beetle cadavers with typical symptoms of infection by entomopathogenic fungi were picked; then, body integuments of every beetle were dissected into several positions as described in protocol step 1.3. As a result, more than 600 fungal isolates were isolated from different body positions. This allows richer fungal candidates for screening pathogens against pine sawyer beetles. Sequencing of the ITS region preliminarily revealed that these isolates can be categorized into 15 fungal genera and 39 species. In addition, there were significant differences in the fungal community composition among geographical populations of M. alternatus adults. The fungal species, Aspergillus austwickii, Akanthomyces attenuatus (syn. Lecanicillium attenuatum), Penicillium citrinum, Scopulariopsis alboflavescens were the dominant species in Sichuan, Zhejiang, Guangdong, Fujian populations, respectively. Under OM and SEM, the asexual reproductive morphology of these four fungi grown on PDA plates was identified based on the macroscopic characteristics, including the color, texture, and sizes of colonies, and microscopic characteristics, including shape, size of conidia, conidiophores, sporangia and hyphae (Figure 2). These observations can be photographed for measuring the shape, size and arrangement of conidia, hyphae, and conidiophores.
The parasitic fungal infection phenotypes on M. alternatus and T. castaneum
Through the induction method (step 3.3), four region-representative fungal species from different geographical populations were confirmed to function in parasitic or non-parasitic modes. As a result, there was no visible infection phenotypes displayed on M. alternatus by P. citrinum, neither on T. castaneum. Conversely, significant infection symptoms were appeared both on M. alternatus (Figure 3) and T. castaneum body surface after infection by A. austwickii, A. attenuatus (syn. L.attenuatum), and S. alboflavescens. Evidence from SEM also well confirmed the consistency of the morphological characteristics of the parasitic fungi on both beetle surfaces. Furthermore, the process of fungal infection on beetles was clearly observed, in which mycelium carrying conidiophores penetrated from the inside of the beetle to the body surface (Figure 4), indicating strong parasitic abilities of these fungi. The above results indicated that the parasitism of these fungi was consistent for both M. alternatus and T. castaneum. In addition, those pictures and data also suggested the spatial preference and localized niches of three fungi on host beetle bodies.
The entomopathogenic activity of parasitic fungal species
T. castaneum has been regarded as a general model organism of Coleopteran species widely, combined with the results above, it has been used as a substitute to assess entomopathogenic activity in this study. The behavioral phenotypes of the three parasitic fungi against M. alternatus and T. castaneum were similar, so the lethal effect on the model beetle would provide valuable information on the entomopathogenic activity against M. alternatus. In a 9-day pilot assay, respectively inoculated with conidial suspension of parasite fungi (A. austwickii, A. attenuatus (syn. L. attenuatum), S. alboflavescens), three groups performed significantly higher mortality on T. castaneum adults than those treated with Tween-80 (control group), and differences were also found between the survival of three groups (Figure 5). These results suggested parasitic fungal species had varying degrees of pathogenicity to model beetles, revealing that these fungi can also exhibit different levels of entomopathogenic activity to M. alternatus adults. Thus, we provide new candidates for the repository of pathogenic fungi.
Multi-gene phylogenetic analysis
The ITS regions of all fungal isolates were subjected to molecular analysis to gain some preliminary insight into the fungal classification. Then, the sequences of the SSU, LSU, tef-1α, rpb2, and β-tubulin region were performed specifically for parasitic entomopathogens. Based on the five independent loci, the accurate taxonomic status of the three parasitic fungi was classified. As shown in Figure 6, the multi-gene phylogenetic treeclearly indicated the genetic distances of the three fungal species from other species in their respective genera, which also matched the morphological characteristics of these fungi.
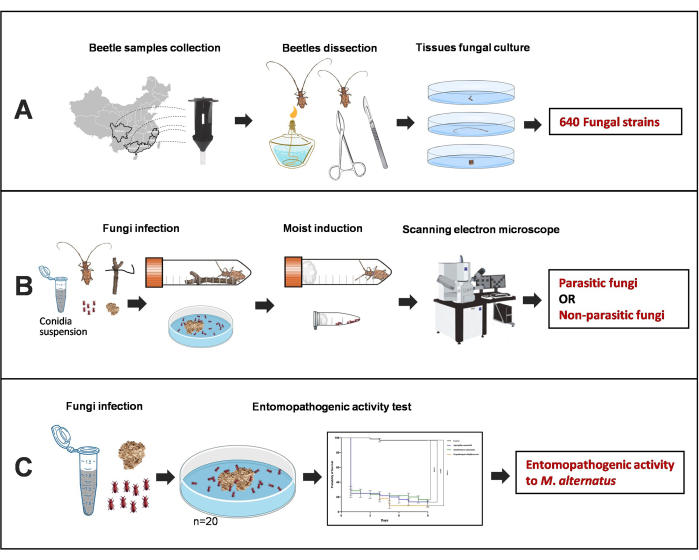
Figure 1: Illustration of screening out entomopathogenic fungi from multiple populations of M. alternatus adults. (A) Fungal isolation from different tissues of M. alternatus with naturally fungal infection. (B) Screening of entomopathogenic fungi. (C) Pathogenicity testing of parasitic entomopathogenic fungi against model beetles. Please click here to view a larger version of this figure.
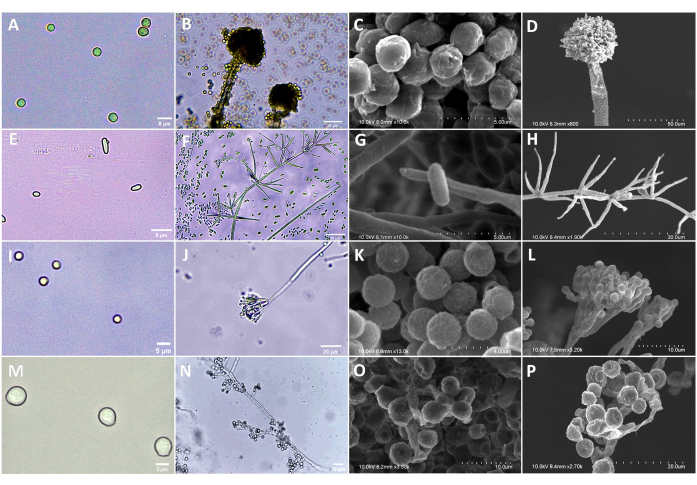
Figure 2: Morphology of A. austwickii, A. attenuatus (syn. L.attenuatum), P. citrinum, S. alboflavescens under OM and SEM. (A) Conidia of A. austwickii (OM). (B) Conidiophores of A. austwickii (OM). (C) Conidia of A. austwickii (SEM). (D) Conidiophores of A. austwickii (SEM). (E) Conidia of A. attenuatus (OM). (F) Conidiophores of A. attenuatus (OM). (G) Conidia of A. attenuatus (SEM). (H) Conidiophores of A. attenuatus (SEM). (I) Conidia of P. citrinum (OM). (J) Conidiophores of P. citrinum (OM). (K) Conidia of P. citrinum (SEM). (L) Conidiophores of P. citrinum (SEM). (M) Conidia of S. alboflavescens (OM). (N) Conidiophores of S. alboflavescens (OM). (O) Conidia of S. alboflavescens (SEM). (P) Conidiophores of S. alboflavescens (SEM). Scale bars are shown in the bottom right corner of each image. Please click here to view a larger version of this figure.

Figure 3: Phenotypes of M. alternatus cadavers infected by A. austwickii, A. attenuatus (syn. L. attenuatum), S. alboflavescens. Beetle cadaver surrounded by mycelium of (A) A. austwickii, (B) A. attenuatus, (C) S. alboflavescens. Please click here to view a larger version of this figure.
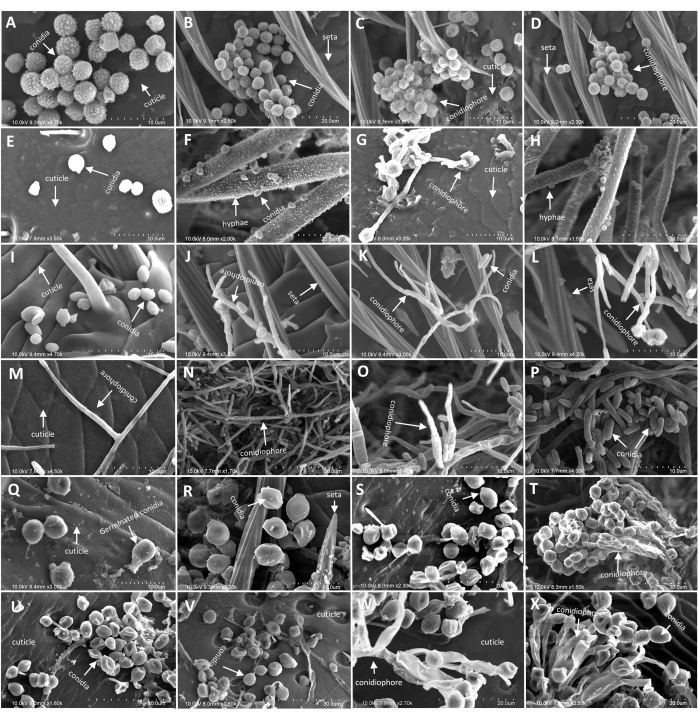
Figure 4: Morphology of M. alternatus and T. castaneum cadaver infected by A. austwickii, A. attenuatus (syn. L.attenuatum), S. alboflavescens under SEM. (A–D) Hyphae and conidiophores of A. austwickii on M. alternatus abdomen. (E–H) Hyphae and conidiophores of A. austwickii on T. castaneum cuticle surface. (I–L) Hyphae and conidiophores of A. attenuatus on M. alternatus antennae. (M–P) Hyphae and conidiophores of A. attenuatus on T. castaneum cuticle surface. (Q–T) Hyphae and conidiophores of S. alboflavescens on M. alternatus head. (U–X) Hyphae and conidiophores of S. alboflavescens on T. castaneum cuticle surface. Scale bars are shown in the bottom right corner of each image. Please click here to view a larger version of this figure.
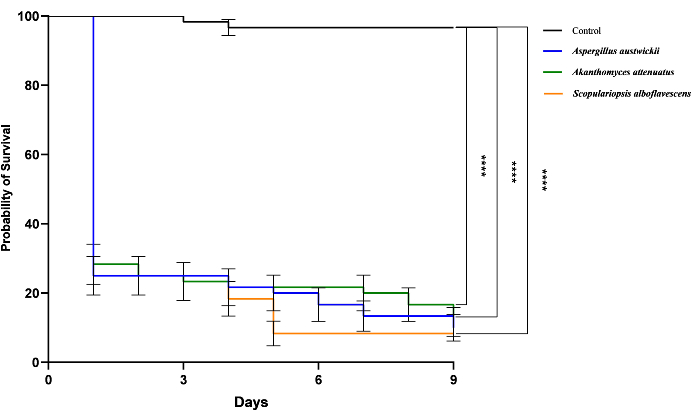
Figure 5: Pathogenicity activities of three parasitic fungi isolated from wild M. alternatus. The survival of T. castaneum beetles infected by three parasitic fungi is shown by Kaplan-Meier curves. Log-rank tests were performed and the significance level were denoted: ns, not significant; ****, P < 0.0001. Please click here to view a larger version of this figure.
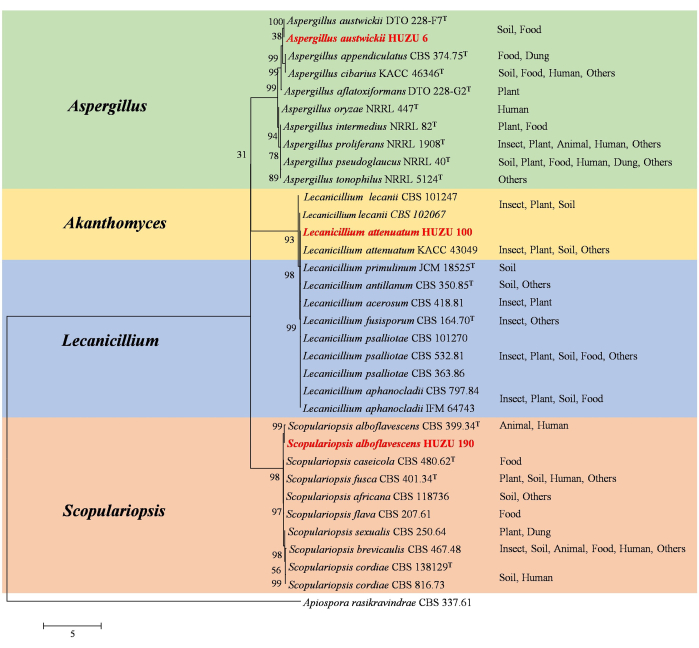
Figure 6: Phylogenetic tree construction of three entomopathogenic fungi inferred from a multi-gene dataset (ITS, LSU, SSU, EF-1α, rpb2, and β-tubulin). The fungal isolates in this study are in red. The nodes indicate supportive values greater than 50%. This figure was modified from Wu et al.43. The habitats where the species were found are indicated behind the species names as Insect, Plant, Soil, Animal, Food, Human, Dung, and others. The T indicates the isolates were from type materials. The GenBank accession numbers of fungal species sequences used for phylogenetic construction see Supplemental Table S1. Please click here to view a larger version of this figure.
| Primer name | Region | Sequence (5'-3') | Bibliography | |
| ITS1 | ITS | TCCGTAGGTGGACCTGCGG | 34 | |
| ITS4 | TCCTCCGCTTATTGATATGC | |||
| NS1 | SSU | GTAGTCATATGCTTGTCTC | 34 | |
| NS4 | CTTCCGTCAATTCCTTTAAG | |||
| LR0R | LSU | ACCCGCTGAACTTAAGC | 37, 38 | |
| LR7 | TACTACCACCAAGATCT | |||
| EF-983 | tef-1α | GCYCCYGGHCAYGGTGAYTTYAT | 39 | |
| EF-2218 | GACTTGACTTCRGTVGTGAC | |||
| RPB2-5’F | rpb2 | CCCATRGCTTGTYYRCCCAT | 40 | |
| RPB2-5’R | GAYGAYMGWGATCAYTTYGG | |||
| TUB1 | β-tubulin | AACATGCGTGAGATTGTAAGT | 41 | |
| TUB22 | TCTGGATGTTGTTGGGAATCC | |||
Table 1: Primer pairs for fungal identification.
Supplemental Table S1: GenBank accession numbers of fungal species sequences used for the phylogenetic construction. Please click here to download this File.
