In the process of assisted reproductive techniques, controlled ovarian hyperstimulation (COH) is a crucial step, leading to significantly elevated estrogen levels in patients by more than 10-20 times compared to natural cycles11. Given this context, we asked whether high serum estrogen concentrations impact embryo implantation during fresh embryo transfer. Based on this embryo implantation model, we studied the effects of various concentrations of E2 and P4 on embryo implantation rates12. After starvation treatment, Ishikawa cells were incubated with E2 (0, 0.1, 1, 10, and 100 nM) or a combination of E2 and P4 (0-0, 100-0, 100-10, and 100-100 nM) for 48 h. Cells were then transferred to gelatin-coated 12-well plates, and 5-15 blastocysts were added to each well. Each group was replicated 3x, and implantation rates were determined after 48 h.
The results demonstrated that near-physiological concentrations of E2 (10 nM) increased embryo implantation rates compared to 0 nM E2. However, ultra-high concentrations of E2 (100 nM) significantly reduced embryo implantation rates. The addition of P4 dose-dependently alleviated the negative effects of ultra-high E2 on implantation (Figure 6). Based on this in vitro model, we illustrate the negative effects of non-physiological ultra-high concentrations of estrogen on embryo implantation and further introduce the application of this model in scientific research.
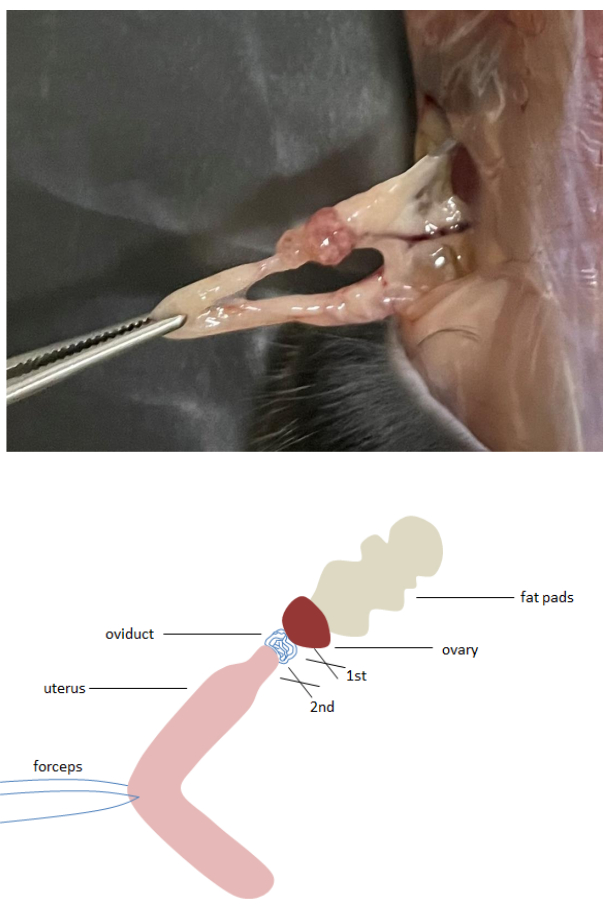
Figure 1: Excision of mouse oviduct. Stretch the oviduct, ovary, and fat pads with forceps, and first cut the oviduct and ovary with scissors, and then excise the oviduct from the uterus. Please click here to view a larger version of this figure.
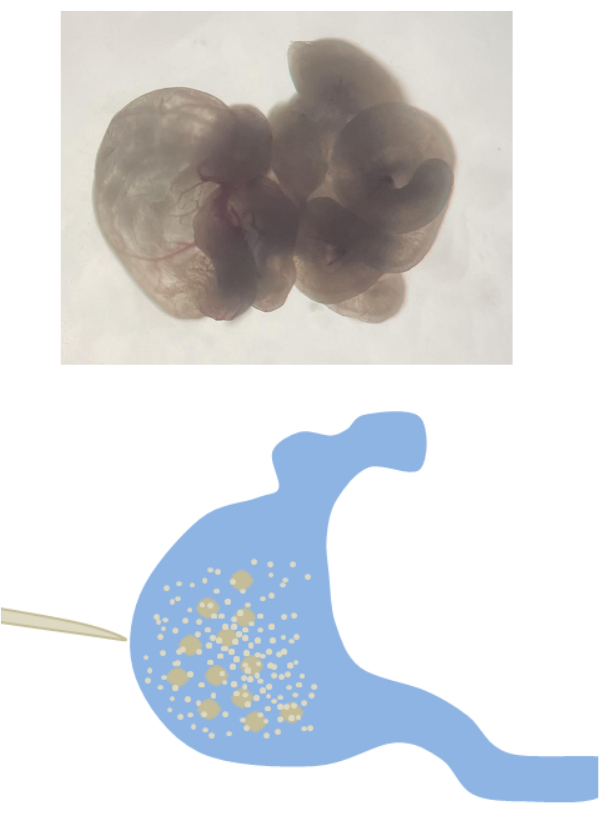
Figure 2: Zygote retrieval. The enlargement ampulla of the fallopian tube is punctured with forceps to release the cumulus-zygote complex. Please click here to view a larger version of this figure.
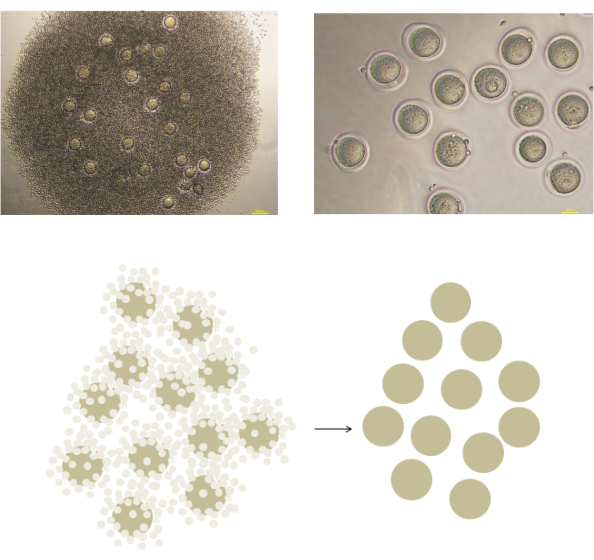
Figure 3: Isolation of Naked zygotes with Hyaluronidase digestion. Using forceps, carefully transfer the cumulus-zygote complex into M2 solution containing 0.5 mg/mL hyaluronidase, and leave it for a few minutes until the cumulus cells detach from the zygotes. Please click here to view a larger version of this figure.

Figure 4: In vitro culture of mouse embryo. Prepare 12 microdroplets of KSOM medium, each with a volume of 20 µL. Cover the microdroplets with 3-5 mL of mineral oil. Transfer the mouse embryos from M2 medium to the upper half of the KSOM microdroplets for thorough washing, place the washed embryos in the lower half, and incubate them at 37 °C with 5% CO2 for 4 days. Please click here to view a larger version of this figure.
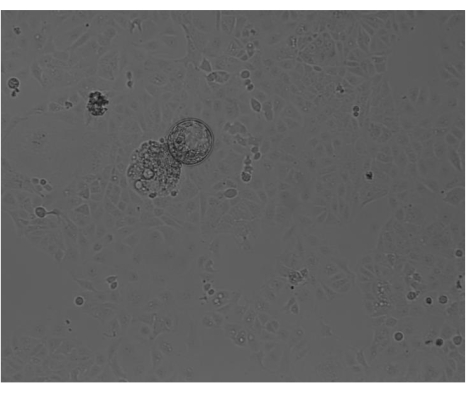
Figure 5: Embryo implantation. Gently place 5-15 fresh blastocysts into each well of the 12-well plate containing Ishikawa cells for co-culture. After 48 h incubation, quickly move the plate 3x. Unattached embryos will exhibit floating or rolling motion over the surface of the Ishikawa cells. Please click here to view a larger version of this figure.
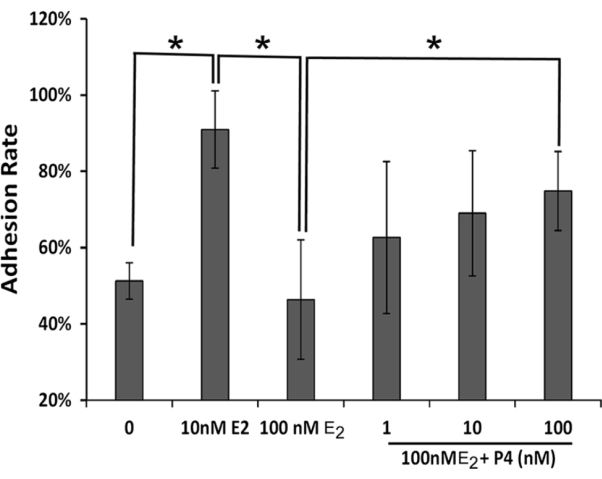
Figure 6: Effect of ultra-high concentration estrogen on the implantation rate of the embryo. The asterisk indicates a statistically significant difference, P < 0.05. Abbreviations: E2 = estradiol; P4 = progesterone. Please click here to view a larger version of this figure.
| 17-β-estradiol | SIGMA | 3301 | Most potent mammalian estrogenic hormone |
| BD Falcon | BD | 353001 | Bacteriological Petri Dishes 35 x 10 mm style w/tight lid, crystal-grade virgin polystyrene, sterile |
| Biosafety Cabinet | ESCO | class  BSC BSC |
Aseptic operations, making culture dishes, aliquoting reagents, etc |
| CO2 Incubator | Thermo | 8000DH | Embryo culture |
| Culture plate | Corning | 3506 | Cell culture |
| DMEM/F12 | Gibco | 1133032 | DMEM/F12 (1x), liquid 1:1,Contains L-glutamine and 15 mM HEPES buffer |
| Fetal Bovine Serum | Gibco | 10099141 | Fetal Bovine Serum, Qualified, Australia Origin |
| Gelatin | SIGMA | G9391 | Type B, powder, BioReagent, suitable for cell culture |
| HCG | Nanjing Aibei | M2520 | Sterilization reagent, intraperitoneal injection, 50 IU/mL |
| Hyaluronidase | SIGMA | V900833 | Reagent grade, powder |
| KSOM | Merck | MR-020P-D | (1x), Powder, w/o Phenol Red, 5 x 10 mL |
| L-glutamine | Gibco | 25030081 | L-glutamine-200 mM (100x), liquid |
| M2 | Merck | MR-015-D | EmbryoMax M2 Medium (1x), Liquid, with Phenol Red |
| Mineral oil | SIGMA | M8410 | Mineral oil is suitable for use as a cover layer to control evaporation and cross-contamination in various molecular biology applications |
| Penicillin-Streptomycin, liquid | Gibco | 15140122 | 10,000 Units penicillin (base) and 10,000 units streptomycin (base), utilizing penicillin G (sodium salt) and streptomycin sulfate in 0.85% saline |
| PMSG | Nanjing Aibei | M2620 | Sterilization reagent ,intraperitoneal injection, 50 IU/mL |
| Progesterone | SIGMA | 5341 | Steroid hormone secreted by the corpus luteum during the latter half of the menstrual cycle |
| Sodium pyruvate | Gibco | 11360070 | Sodium pyruvate is commonly added to cell culture media as a carbon source in addition to glucose |
| Stereomicroscope | Olympus | SZX7 | Embryo retrieval and observation of embryo development |
