Primary rod shaped cardiomyocytes attach on laminin matrices and spread and differentiate into confluent cultures (Figure 1A and its inset). These cells were further characterized on the basis of sarcomeric actin and SERCA2 expression (Figures 1B and 1C). Rat cardiomyocytes are highly susceptible to chlorine toxicity as 15 min exposure to 100 ppm chlorine caused extensive cell rounding and death in submerged cultures and disruption of confluent layers on cells grown on laminin coated membranes (Figure 1D). There was also enhanced apoptotic cell death as indicated by caspase 3/7 release in cardiomyocytes grown on inserts (Figure 1E).
Exposure of differentiated human airway epithelium (Figure 2, inset to panel 1 showing a cross section of cell culture inserts with columnar, ciliated and goblet cells) to chlorine caused sloughing and lifting of cell membranes at both low (100 ppm) and high (300 ppm) concentrations (Figure 2 panel A2 and A3). Damage by low chlorine concentrations was quickly reversed (Figure 2 panel A5), however, cells exposed to higher chlorine concentrations had delayed or no repair ability (Figure 2 panel A6) as shown by visual inspection as well as cell proliferation assessment by Ki-67 staining (Figure 2 panel A9). Trans epithelial electrical resistance, TER measurements and caspase activity further confirmed these results and provide evidence for loss of membrane integrity and apoptotic cell death upon chlorine exposure (Figure 2, panel B and C). Thus our studies describe the development of an in vitro Cl2 exposure system that causes loss of membrane integrity and death of airway epithelium and cardiomyocytes. This effect may not be due to non-physiological pH changes in the cardiomyocytes as the pH of the media after exposure to chlorine was maintained at ~7.4 as measured by using a pH meter in the collected media postexposure.
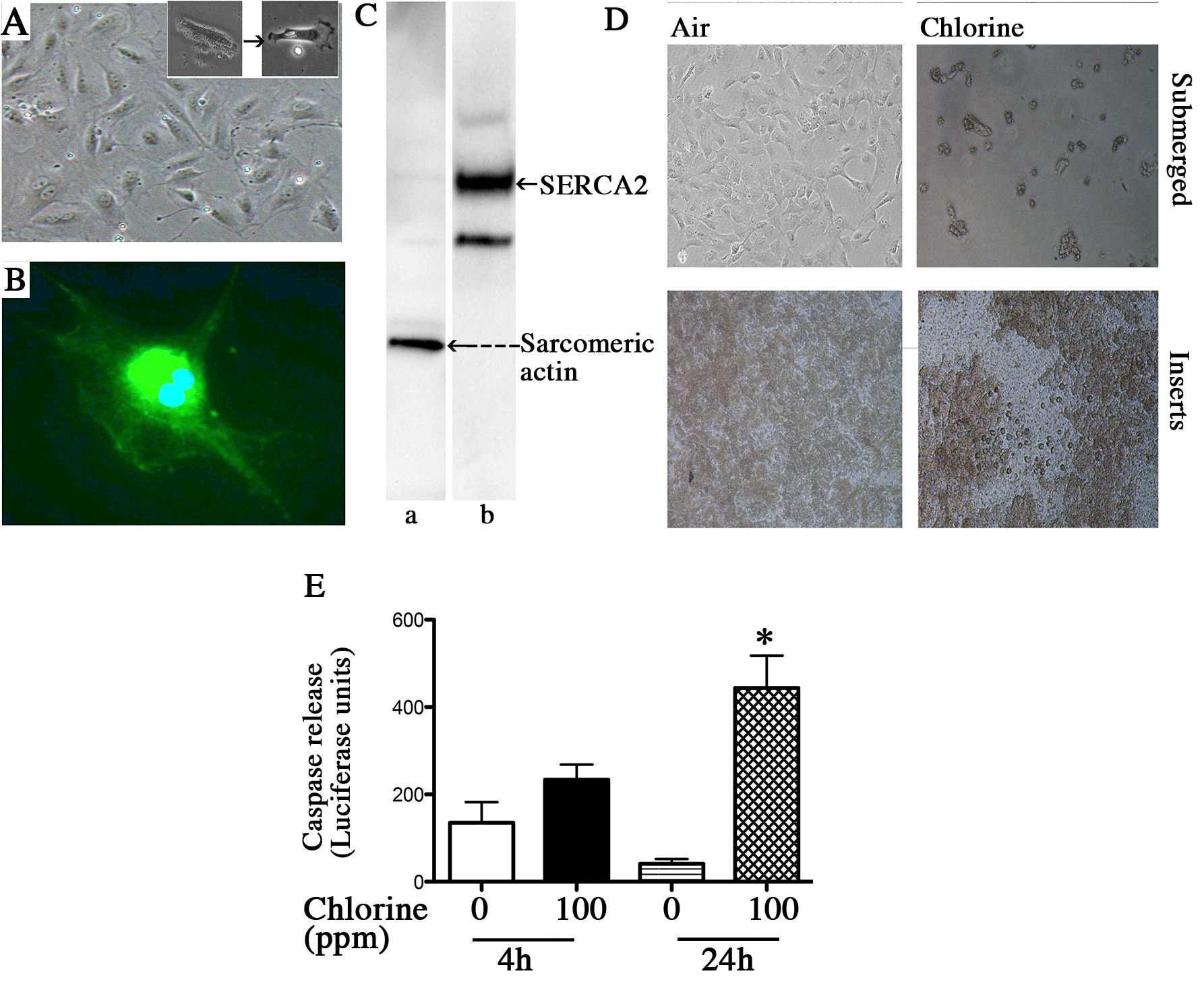
Figure 1. Rat cardiomyocyte isolation and exposure to chlorine. Rat cardiomyocytes were isolated as described in the protocol and plated on laminin-coated plastic dishes (panel A, a representative light microscopic image) or laminin coated inserts. Inset to panel A shows a rod shaped cardiomyocyte spreading and differentiating. The cardiomyocytes were also characterized based on the sarcomeric actin expression (panel B showing a representative image detected by immunofluorescence and lane a panel C showing a representative western blot scan) and abundant SERCA2 expression (panel C lane b). Chlorine exposure (100 ppm 15 min) caused extensive cell death in submerged cultures as well as disruption of the cell monolayer on the inserts (panel D showing the representative photomicrographs). Caspase 3/7 release (panel E) in the supernatant media of cardiomyocytes grown on inserts, at 4 hr and 24 hr post chlorine exposure was also measured as described in the text. Values shown are mean ± SEM and * indicates significant (p < 0.05) difference from 0 ppm control.
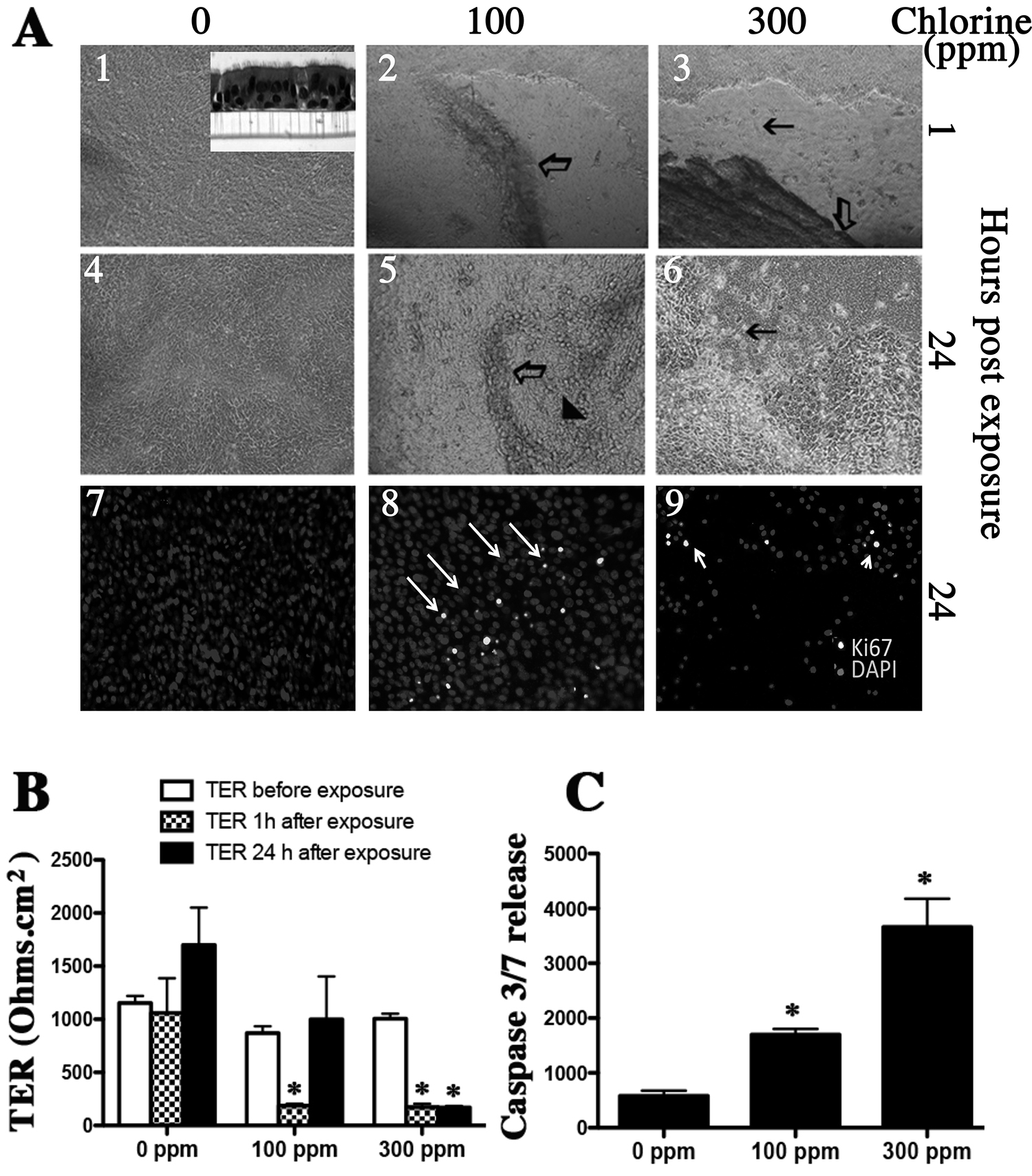
Figure 2. Effect of chlorine exposure on differentiated human airway epithelial air-liquid interface (ALI) cultures. Human airway epithelial basal cells were cultured on collagen coated snapwells. After day 5 the apical media was removed. Differentiated cultures (consisting of basal, ciliated, columnar, and goblet cells as shown in the inset in top left panel of panel A) were exposed to chlorine (100 or 300 ppm) for 30 min. The TER was measured and media was changed and cells incubated for 24 hr. At 24 hr TER was measured again and apical media was collected for caspase release measurement and the cell membranes were fixed for immunohistochemistry. The open arrow in panel A parts 2 and 3 shows sloughed off epithelial layer and black arrows show empty spaces on the insert. The arrowhead in panel A part 5 shows regenerated epithelium. Parts 7, 8, and 9 show cellular proliferation as assessed by Ki-67 immunostaining. Values shown are mean ± SEM and * indicates significant (p < 0.05) difference from 0 ppm control.
