1. Cell Preparation
- Grow overnight cultures of the desired strain carrying the sticky fliC allele 15,22 in Tryptone Broth (TB, 1% Peptone, 0.5% NaCl) followed by inoculation at 1:100 dilution in 10 mL fresh TB. Grow the culture at 33 °C in a shaker incubator until OD600 = 0.5.
- Pellet the cells at 1,500 x g for 5 – 7 min and re-disperse the pellet vigorously in 10 mL of filter sterilized motility buffer (MB; 10 mM phosphate buffer: 0.05-0.06 M NaCl, 10-4 M EDTA, 1 µM methionine, pH 7.0).
- Repeat step 1.2 two more times and re-disperse the final pellet in 1 mL MB.
- Shear the suspension by passing back and forth ~ 75 times between two syringes with 21 to 23 gauge adapters connected by polyethylene tubing (7 – 12 cm long, 0.58 mm inner diameter). Limit the total time for shearing to 30 – 45 s.
- Centrifuge the sheared cells at 1,500 x g for 5-7 min and re-disperse the pellet in 100 – 500 µL of MB.
2. Slide Preparation
- Prepare an imaging chamber by sandwiching two double-sided adhesive tapes between a cover-slip and a microscope slide. For chemotaxis assays, employ any microfluidic chamber that enables the exchange of MB and chemical stimulants.
- Add 0.01% poly-L-lysine solution in the chamber and after 5 min gently rinse the surfaces with MB (80 – 100 µL).
- Add 40 µL of the cell suspension into the chamber and allow sufficient time for attachment to the glass surface (7 – 8 min). Flow out unstuck cells by adding 100 µL MB on one side of the chamber, while wicking the solution with a filter paper from the other side.
- Add 10 – 15 µL of latex beads into the chamber and allow the beads adequate time to settle and attach to the cells (7 – 8 min). Gently rinse with 100 µL of MB, as described in step 2.3, to remove unstuck beads. Use a range of bead-sizes for the experiments so long as a good contrast is available.
3. Bead Tracking
- Place the sample on a microscope stage and scan the surface for beads attached to motors. Use a 40X phase objective to make observations although phase microscopy is not necessary. Alternatively, employ bright-field imaging so long as sufficient contrast is maintained to clearly distinguish a bright bead on a dark background.
- Once a bead has been selected, move the stage laterally to position the bead in a pre-determined corner as shown in Figure 1B. Position beads at the same corner to ensure that the direction of rotation of the bead is correctly known. The ideal bead trajectory is approximately circular but elliptical trajectories are admissible.
- Maintain the sampling frequency higher than twice the rotational frequency of the motor to avoid errors associated with aliasing. In this work, use a motor that was rotating at 50 Hz and sample at frequencies that were 10 times higher (500 Hz), to obtain a smooth signal.
4. Data Analysis
- Center and scale the PMT output voltages and correct ellipticity in the trajectories with affine transformations if needed 23. Use a power-spectrum analysis to determine rotation rates 17.
- Determine polar angles, θ(t) = atan (y(t)/x(t)). Determine the variations in motor speeds and switching over time by calculating ω
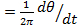 14.
14. - Employ a median filter to smooth the motor speed data. A filter window over two full rotations is recommended 23,24.
The photomultiplier setup is shown in Figure 1A. It is important that the PMTs have high sensitivities over the range of wavelengths scattered by the beads of interest. The PMTs employed here operate in the visible and near-infrared ranges, and were able to detect light scattered by beads illuminated by a halogen light source. The optimum lighting conditions and supply voltages will vary from one setup to another. For the setup used in this work, a PMT gain ~ 104 – 105 proved adequate. Each photomultiplier was covered except for a 3 x 1 mm slit positioned in front of the photomultiplier. The slits limit the region in the cell-sample from which light can enter the photomultipliers, and the two slits are orthogonal to one another. When a rotating bead is positioned at the correct location (Figure 1B), the amount of light entering the photomultiplier increases as the bead comes in the view and decreases as its circular path takes it away from the view. The frequencies of the sinusoidal PMT voltage outputs indicate the speed of rotation and the phase differences between the two signals indicate the direction of rotation. The use of an oscilloscope to display the PMT outputs enables visualization of bead trajectories in real-time.
The time-varying PMT signals, y(t) and x(t), from a representative motor are shown in Figure 2A. The orthogonality of the two slits introduces a phase lag between the two signals. The signal amplitudes depend on the signal-to-noise ratio as well as the eccentricity of rotation. The corresponding trajectories of the bead are indicated in Figure 2B.
A histogram of the speeds measured from a representative motor in a cheY– deleted strain is shown in Figure 3A. The deletion ensures that flagellar motors cannot switch and rotate exclusively counterclockwise (CCW). The average speed from this particular motor was 60 Hz and the measured speeds are consistent with previous reports for motors tethered with 1 µm beads 25. The bead was first positioned at the lower right corner, as seen in the schematic in Figure 1B. The corresponding angular speed is shown in Figure 3B (top panel). Positioning the bead to the adjacent lower left corner resulted in inversion of the sign on the motor speeds (bottom panel). Thus, moving the bead to an adjacent corner will change the observed direction of motor rotation. In this regard, diagonally opposite corners are identical. It is therefore crucial to know the location of the bead during measurements to correctly determine the switching dynamics. Figure 3C shows repeated transitions of a wild-type motor between the two directions of rotation.
Custom codes for data-acquisition software were adapted from prior work to record the data on a computer 15. The PMT output was AC-coupled and low-pass filtered with a cutoff frequency of 100 Hz. Real-time tracking was enabled by connecting the filtered outputs to an oscilloscope.
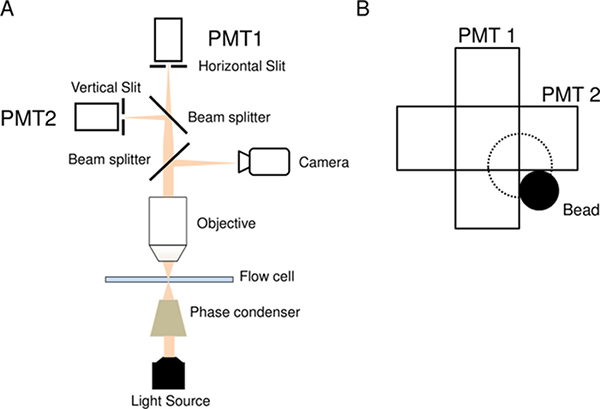
Figure 1: Bead-tracker Setup. A) Schematic of the PMT-based tracking setup. B) The ideal position of the bead (black sphere) relative to the two orthogonal slits. The trajectory is indicated by the dotted lines. The eccentricity e is the radius of the dotted circle. Please click here to view a larger version of this figure.
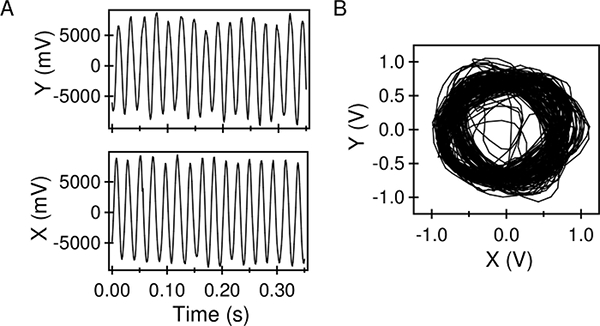
Figure 2: PMT Outputs. A) Low-pass filtered outputs from the two photomultipliers, after centering/scaling. B) The bead trajectories obtained from the PMT data, sampled over 3 s. Please click here to view a larger version of this figure.
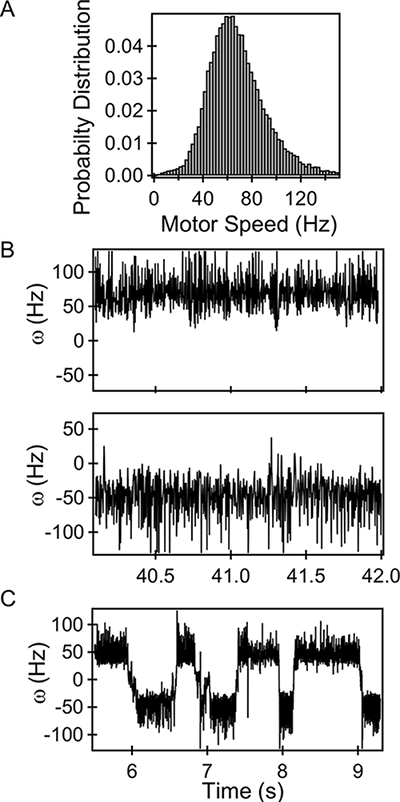
Figure 3: Bead Trajectories. A) Histogram of CCW-only speeds of a representative motor. B) Rotational speeds of a CCW-only motor imaged at lower right corner (top panel). Rotational speeds of the same motor when positioned at the lower left corner (bottom panel). C) Switching in a representative wild-type motor. Please click here to view a larger version of this figure.
