1. Cloning Considerations
- Commercially prepared oligonucleotides representing the TMD of interest flanked by NheI and BamHI restriction sites and 5′-phosphorylated can be ligated into pTox7 (modified in our laboratory by insertion of one base pair directly after the BamHI restriction site14) (Figure 3) digested sequentially with BamHI and NheI. An example oligonucleotide is shown below:
5’ctagcTMDSEQUENCEg3′
3′ gTMDSEQUENCEcctag5′
The TMD sequence should be 12-24 residues (shorter sequences will presumably be elongated by vector encoded hydrophobic residues). In order to investigate the interface, four variants of the TMD design should be created where sequential residue insertions and concomitant residue deletions result in rotation of the TMD relative to ToxR.15,16 Finally, the arabinose concentration should be varied between 0.001 and 0.01% (w/v) to identify the concentration where maximum differences in β-galactosidase signals between different TMD sequences are observed; testing different expression levels is recommended to identify conditions under which different affinities can be distinguished best. In addition to arabinose and antibiotics, 0.4 mM IPTG can be used to enhance differences of affinities between different TMDs. The ToxR measurement should be performed at least in quadruplicate. The whole procedure should be repeated at least three times with different plasmid transformations.
2. Growth of Bacterial Cultures
- Gently thaw FHK12 competent cells (200 μl) on ice and transfer into a 15 ml culture tube. Add plasmid DNA (200 ng) and incubate the cells on ice for 30 min.
- Heat-shock cells by incubation for 90 s at 42 °C, followed by incubation on ice for 2 min.
- Add SOC media (800 μl) and incubate the samples at 37 °C with shaking (300 rpm) for one h.
- Inoculate 5 ml LB media with chloramphenicol (30 μg/ml) and arabinose (0.0025% w/v) with 50 μl of the transformation mixture in 15 ml culture tubes in triplicate. Incubate samples at 37 °C with shaking (300 rpm) for 20 h. (Alternatively 5 μl of culture can be used to inoculate 100 μl of medium in a 96-well plate. This method is useful when dealing with large numbers of samples, although errors will be slightly higher. In order to avoid evaporation, which would lead to error, fill the outermost wells with media, but do not use them for samples. Finally, double-wrap the joint between the lid and the plate with parafilm).
3. Measurement of β-galactosidase Activity
- Preheat the plate-reader to 28 °C.
- Transfer the Z-buffer into a reservoir with a large pipette tip, making sure to only take up the upper (aqueous) layer. Transfer 100 μl of freshly prepared Z-buffer/chloroform to the wells of a 96-well plate. Transfer 5 μl of each culture into the wells of the plate in quadruplicate. Omit the culture from four wells which will serve as the blank.
- Measure the OD595 of the plate to determine cell density.
- Add 50 μl of Z-buffer/SDS to all wells of the plate. Shake the plate in the plate-reader for 10 min to lyze the cells. Make sure the cell suspensions are clear after lysis and repeat the shaking step if required. Incomplete lysis suggests the Z-buffer/chloroform was not freshly prepared.
- Add 50 μl freshly prepared Z-buffer/ONPG to all wells and return the plate to the plate reader and measure OD405 every 30 s for 20 min.
- Calculate β-galactosidase activity using the following equation (remembering to subtract the blank). The ratio of OD405/min should be calculated using all data points in the OD405 range 0.0 to 1.0 using a linear model fit.
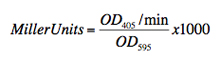
Miller units differ sometimes when recorded on different days. Therefore, a reference construct like GpA should be measured in each test. Its values can be used for normalization of ToxR values.
4. Control for Protein Expression
- Perform Western blotting to verify even protein expression between constructs. Combine 50 μl of the triplicate cultures and centrifuge (2000 rpm, 4 min) in a microcentrifuge. Remove the supernatant by pipetting and resuspend the residual pellet in 2 x sample loading buffer.
- Load 7.5 μl on a standard 8% gel and carry out electrophoreses at 125 V for 1 hour 5 min. After transfer, incubate with anti-MBP HRP-conjugated antibody and visualize; the chimeric protein is observed at approximately 70 kDa with some degradation products sometimes seen around 48 kDa. Endogenous MBP is also observed at 45 kDa (see Figure 5).
5. Control for Proper Membrane Insertion
A cell line deficient in maltose binding protein is used to assess proper membrane insertion of the chimeric TMD construct. When grown on minimal media with maltose as the sole carbon source, only cells expressing a membrane-integral expression product with maltose binding protein correctly located to the periplasm are able to grow.
- Transform PD28 cells (as described for FHK12 cells) and inoculate 2 ml of LB medium. Grow the cells at 37 °C with shaking (300 rpm) overnight.
- Pellet the cells by centrifugation at 3500 rpm, 10 min, 4 °C and wash by resuspension in PBS (2 ml) by gentle pipetting with a large tip or gentle vortexing. Pellet the cells (as above), wash with PBS for a second time, pellet and finally resuspend in PBS (1 ml).
- Use 25 μl of resuspended cells to inoculate 5 ml minimal media in triplicate and incubate at 37 °C with shaking (300 rpm). Take OD595 readings between 15-25 h, approximately every 2 hours by transferring 200 μl of each sample into a 96-well plate and reading using the plate-reader.
6. Representative Results:
An example of the use of the ToxR transcriptional reporter assay to analyze the oligomerization propensity of transmembrane domains in shown in Figure 4. Previously we have investigated the oligomerization of transmembrane domains from the multispanning membrane-integral protein latent membrane protein-1 (LMP-1) by various techniques, including ToxR.14 Transmembrane domain five (TM5) was shown to exhibit a strong propensity to oligomerize; this is demonstrated by high Miller Units, comparable to the positive control, GpA, a well-established dimerizing sequence. A deleterious mutation in TM5, D150A, reduces the ability of the sequence to oligomerize. LMP-1 TM1 does not significantly oligomerize and exhibits a very low Miller Unit signal, just above the signal for blank, non-transformed FHK12 cells.
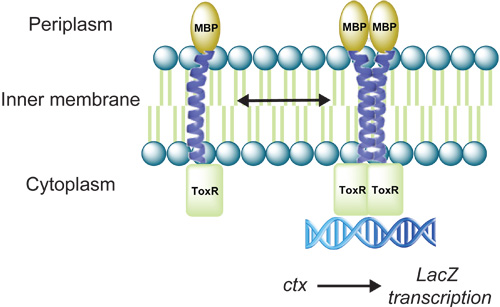
Figure 1. Cartoon depicting the ToxR reporter assay. Transmembrane domain (TMD) driven oligomerization results in dimerization of ToxR and activation of LacZ transcription. The gene product of LacZ, β-galactosidase can be quantified as a measure of the propensity of a TMD to oligomerize.
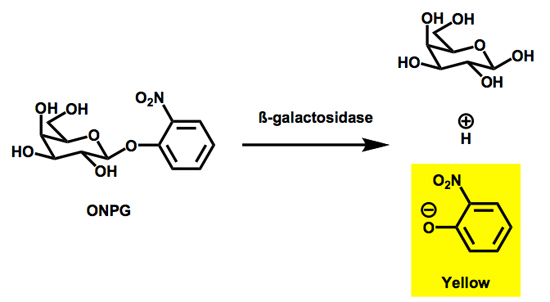
Figure 2. The hydrolytic cleavage of ONPG by β-galactosidase results in the production of the light absorbing species o-nitrophenolate (ONP).
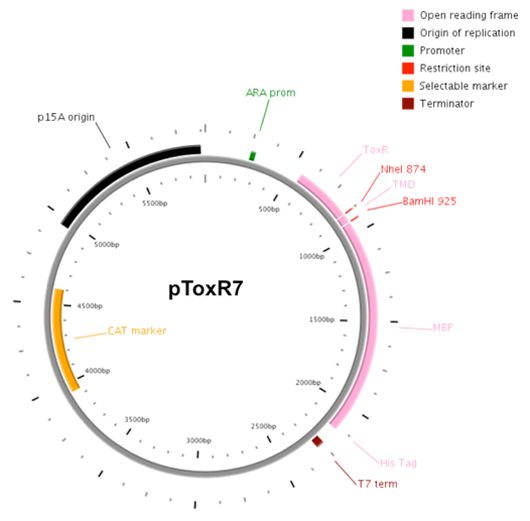
Figure 3. Plasmid map of pToxR7.
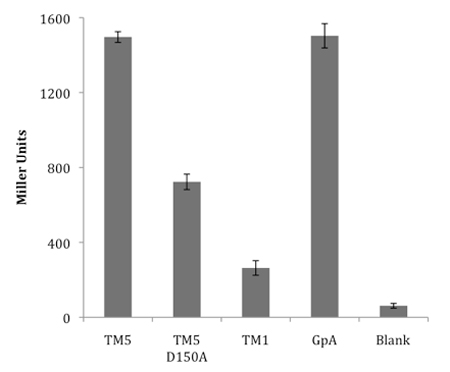
Figure 4. Representative ToxR transcriptional reporter assay analysing the oligomerization propensity of latent membrane protein-1 transmembrane domains. Transmembrane domain 5 (TM5) oligomerizes strongly, whilst transmembrane domain 1 (TM1) exhibits only a weak interaction. Mutation D150A in TM5 significantly reduces its ability to oligomerize. GpA is included as a positive control sequence for strong dimerization. Blank represents untransformed FHK12 cells.
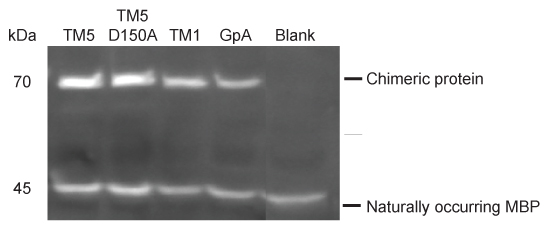
Figure 5. Western blot for protein expression.
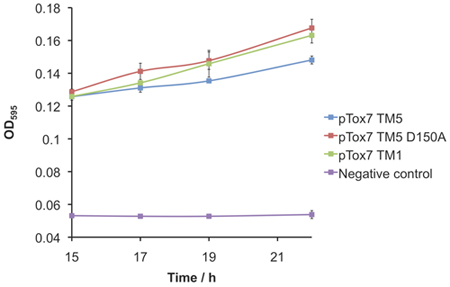
Figure 6. PD28 complementation assay to control for correct membrane insertion to the periplasm. Negative control represents a construct deficient in maltose binding protein.
