1. Transfection with Target ID Library and Selection for Stable Cell Lines
1. Zeocin Kill Curve
Zeocin is used to select for stably transfected cells. However, excess zeocin causes undesired phenotypic responses in most cell types. Therefore, a kill curve analysis must be performed to establish the minimum lethal dose.
- Plate 1.6 x 104 cells into wells of a 96-well plate in 120 μl of media.
- The next day add zeocin in increasing concentrations ranging from 50 μg/ml to 1 mg/ml to the appropriate wells.
- Examine viability every 2 days.
- Replace media containing zeocin every 3 days. The minimum concentration of selection reagent that causes complete cell death after the desired time should be used for that cell type and experiment. Our results show that 500 μg/ml zeocin is optimum for A549, HeLa, and MCF7 cells.
2. Library Transfection via Nucleofection and Selection
- Select a cell line that either does not express or expresses low levels of your miRNA of interest. The miRNA will be introduced in Section B for target selection after stable expression of the Target ID Library is achieved.
- Culture/expand cells. We have obtained excellent results with 2 x 107 cells per library transfection.
- Trypsinize cells that are at >80% confluency, and transfer 2 x 107 cells to a 15 ml sterile screw topped tubes.
- Pellet trypsinized cells at 200 x g for 5 min.
- Remove medium and wash cell pellet with HBSS or 1X PBS.
- Centrifuge at 200 x g for 5 min and aspirate wash.
- Repeat wash step of cell pellet.
- Pre-warm 6-well plates with 2 ml of complete medium at 37 °C.
- Add 2 μg of Target ID Library (not to exceed 10 μl) per 0.5 ml tube for each transfection.
- Resuspend cells in the 15 ml tube from above with 100 μl of Amaxa Nucleofection Solution (cell specific) per 2 x 106 cells. For example, for 10 Nucleofections add 1 ml of reagent.
- One reaction at a time, add 100 μl of cells to the 2 μg of plasmid. Mix with pipette.
- Transfer mixture to a Nucleofector cuvette.
- Insert cuvette into Nucleofector instrument and run optimized program appropriate for the cell line (High Efficiency preferred over Cell viability).
- Fill transfer pipette with pre-warmed medium. Take up cells in same pipette and transfer to 6-well plate. Repeat for each Nucleofection, one per well.
- Return to growth chamber for overnight incubation.
- The next day, replace medium and allow cells to recover for 3-5 days.
- Replace medium with complete medium containing the appropriate level of zeocin, as determined from the kill curve analysis.
- Monitor cells for zeocin selection (cells dying).
- Replace medium with zeocin every 2-3 days.
- Once confluent in 6-well plate, passage, pool and expand cells in larger flasks.
- Length of time for expansion of cells is user and cell line dependent, but it is highly recommended to expand zeocin resistant cells to generate cryo-stocks for future screening (~2-3 weeks; cell line dependent).
2. Transfect Library Cells with miRNA-Expression Construct, Select for Stable Cell Line, and Select miRNA Targets
Note: Zeocin selection is no longer required or desired. Exposure of cells to zeocin during miRNA expression and ganciclovir (target) selection may result in loss of miRNA targets.
3. Puromycin, G418, and Ganciclovir Kill Curves
- Perform a kill curve for ganciclovir with cells stably expressing the Target ID Library and with puromycin or G418 for wild type cells.
- Plate 1.6 x 104 cells into wells of a 96-well plate with 120 μl fresh media. The next day add from 0.1 to 10 μg/ml of puromycin/G418, or 2 to 32 μM ganciclovir to selected wells.
- Examine viability every 2 days. Replace the media containing selection reagent every 3 days. The minimum concentration of selection reagent that causes complete cell death* after the desired time, should be used for that cell type and experiment. Our results show that 0.25 to 1 μg/ml puromycin is optimum for A549, HeLa, and MCF7 cells, 0.3 μg/ml G418 for MCF7 cells and 8-16 μM ganciclovir are optimum for A549, HeLa, and MCF7 cells.
*Note: With ganciclovir selection, slow or no cell growth may be observed without complete cell death. Observe cells over several days’ treatment to verify they are not actively dividing. If this is the case then phenol red in the medium remains red. It may also be necessary to perform a kill curve analysis of the Target ID library transfected cells if a change in susceptibility is observed. However, this will need to be determined on a cell type specific basis.
4. miRNA Transfection via Nucleofection and Target Selection
- Grow / expand Target ID Library cells to attain 2 x 107 cells, and trypsinize when cells are at >80% confluency.
- Transfer 2 x 107 cells to a 15 ml sterile screw topped tube and pellet at 200 x g for 5 minutes.
- Remove medium and wash cell pellet with HBSS or 1X PBS.
- Centrifuge at 200 x g for 5 min and aspirate wash.
- Repeat wash step of cell pellet.
- Pre-warm 6-well plates with 2 ml of complete medium per well at 37 °C.
- Add 2 μg of miRNA expression plasmid (not to exceed 10 μl) per 0.5 ml tube for each transfection. Origene MicroRNA Expression Plasmids (Neomycin – G418 selection) or self-cloned miRNA genes in pBABE-Puro (Plasmid 1764; Addgene) have been used successfully. For the latter, the miRNA hairpin with ~ 200 bp on either side was PCR cloned from human DNA.
- Resuspend cells in the 15 ml tube from above with 100 μl of Amaxa Nucleofection Solution (cell specific) per 2 x 106 cells. For example: For 10 Nucleofections, add 1 ml of reagent.
- One reaction at a time, add 100 μl of cells to the 2 μg of plasmid. Mix with pipette.
- Transfer mixture to a Nucleofector cuvette.
- Insert cuvette into Nucleofector instrument and run optimized program appropriate for the cell line (High efficiency preferred over cell viability).
- Fill transfer pipette with pre-warmed medium. Take up cells in same pipette and transfer to 6-well plate. Repeat for each Nucleofection, one per well.
- Return to growth chamber for overnight incubation.
- Replace medium and allow cells to recover for 3-5 days.
- Replace medium with complete medium containing the appropriate level of puromycin or G418 as determined from the kill curve test performed before transfection with the miRNA construct.
- Monitor cells for puromycin / G418 selection (cells dying).
- Replace medium with puromycin / G418 every 2-3 days.
- Once confluent in 6-well plate, passage, pool and expand cells in larger flasks.
- Length of time for expansion of cells is user dependent, but it is highly recommended to expand puromycin / G418 resistant cells to generate cryo-stocks for future screening (~2-3 weeks; cell line dependent).
- Replace medium with the appropriate levels of ganciclovir (GCV) and puromycin / G418, as determined from the kill curve test performed before transfection with the miRNA construct.
- Monitor cells for selection (cells dying, miRNA targeting and knockdown of TK-ZEO).
- Expand cells in the presence of GCV and puromycin /G418.
- Prepare Genomic DNA from the GCV selected cells.
- PCR amplify inserts with kit primers (see PCR Amplification).
- Clone into the TOPOTA vector (Invitrogen) for standard sequencing or submit PCR product for deep sequencing.
3. PCR-Amplify Selected Library Inserts and Sequence
Note: This procedure is performed to PCR-amplify the library targets that have survived the ganciclovir selection.
5. Collect ganciclovir selected cells to prepare chromosomal DNA using a GenElute Mammalian Genomic DNA Miniprep Kit (Catalog Number G1N10) or equivalent. We recommend preparing DNA from the parent cell line that does not contain Target ID Library to use as a negative control for comparison with DNA from target-selected cells in PCR.
- PCR Amplify the genomic DNA with Amplification Primer 1 and Amplification Primer 2. Successful amplification has been obtained with JumpStart REDTaq ReadyMix Reaction Mix – for PCR (Catalog Number P0982). Optimization of the conditions may be necessary if another polymerase is used. Refer to Table 1 for a sample PCR setup and Table 2 for PCR cycling conditions.
- Resolve 2-5 μl of the PCR product on a 1% agarose gel. The expected product should be a smear with some visible DNA banding. Compare to control PCR product of genomic DNA from cells without the Target ID Library.
- Proceed with cloning and/or deep sequencing of PCR product. If no amplification product is observed or is identical to control DNA, optimize the PCR amplification conditions (i.e., primer concentration, annealing temperature, and cycles).
6. Cloning and Sequencing
Note: We highly recommend cloning the PCR products and then performing standard sequencing from at least 96 of the clones using the Amplification Primers provided in the kit. This procedure has been performed successfully using 96-well overnight cultures and plasmid purification systems. Even if deep sequencing is desired, preliminary results from cloning and standard sequencing can be used as a quality check to determine whether the extra expense of deep sequencing is warranted.
- Follow TA cloning kit manufacture’s protocol for cloning PCR amplification products (above).
- Transform clones into competent bacterial cells and select overnight on antibiotic containing medium.
- Isolate and grow individual colonies in liquid culture containing appropriate antibiotic.
- Purify plasmid DNA. 6.5. Perform sequencing reactions with Amplification Primers.
- Identify gene targets by BLAST alignment of sequences with human transcriptome (for known transcripts) and human genome (for novel transcripts).*
*Cloning troubleshooting: If a high number of inserts from sequencing are plasmid sequence, optimize the cloning/sequencing conditions as such:
- PCR Amplification product (insert) quality and quantity
- Ligation reaction
- Plasmid preparation (quality and quantity)
- Sequencing reaction conditions
4. Representative Results
Library screen for miR-373 targets
To evaluate performance of the Mission Target ID Library, miR-373 targets were selected from MCF-7 library expressing cells. MCF-7 was chosen because it expresses little or no detectable miR-373 (data not shown). miR-373 was chosen for its biological interest. miR-373 expression promotes tumor invasion and metastasis in MCF-7, normally a non-metastatic cell line[2]. Furthermore, miR-373 orthologues from the mouse miR-290 cluster are involved in mouse embryonic stem cell maintenance[3], and the miR-373 family member, miR-372, promotes fibroblast reprogramming to induced pluripotent stem cells[4]. Finally, we used zinc finger nucleases to insert a PGK promoter-miR-373 expression construct into the AAVS1 site in MCF-7 cells, and generated a list of potential miR-373 targets by RNA microarray analysis (data not shown).
The Target ID Library was transfected into MCF-7 cells, and a stable population of cells was selected and amplified in zeocin-containing medium. The resulting cells (MCF-7 Library) were stably transfected with a miR-373 expressing construct and selected in ganciclovir-containing medium to enrich for cells expressing miR-373 targets. We observed that the negative control MCF-7 Library cells (without miR-373) did not detach and become rounded as expected for dead cells. However, they did stop growing, which was readily detected because the phenol red-containing medium did not turn orange-yellow as observed for cells expressing miR-373 (Fig 5). Cells were expanded in ganciclovir-containing medium, and target sequences were isolated by PCR with primers flanking the Target ID Library cDNA inserts and DNA prepared from the surviving cells. PCR products were Illumina sequenced.
As shown in Table 4, we obtained 17,740,719 cDNA reads, which mapped to 11,076 unique genes. Of these unique genes, 2,898 were detected with more than 40 reads, and therefore were considered reliable hits. The 13,106,469 vector reads were expected because the PCR primers used to amplify cDNA inserts are 40 and 300 bases away from the insertion site. As such, it is expected that most resulting sequences would be from the vector unlike more traditional deep sequencing of genomic material. Fourteen percent did not map to either vector or cDNA, but did align with NCBI’s non-redundant nucleotide database (i.e., with NR hit), and only 1% had no cDNA insert.
An initial comparison found that 10 of the unique genes identified with the Target ID Library are also in the list of previously identified miR-373 targets in TarBase (Table 5). These 10 miR-373 targets were previously identified by microarray with RNA from HeLa cells transiently transfected with a synthetic miR-373 mimic[5]. Furthermore, we detected these same 10 genes down-regulated in MCF-7 cells expressing miR-373 from the AAVS1 site (data not shown). Therefore, these 10 are likely valid targets of miR-373. Work is in progress to characterize the list of potential targets and attempt to validate selected hits experimentally by RT-qPCR, Western blot, and luciferase reporter assay.
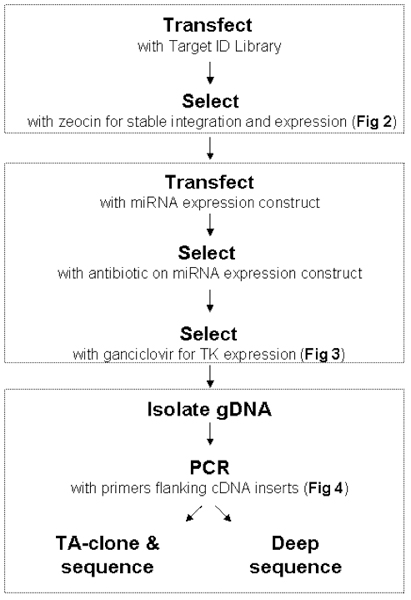
Figure 1. Workflow for Target ID Library. Sections A-C: Refer to sections in the Procedure. Each step is illustrated and described in detail in Figs 2 and 3.
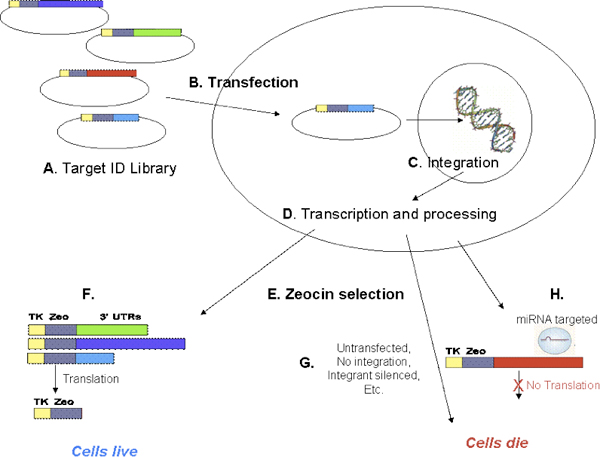
Figure 2. Transfection and Zeocin selection. The Target ID Library is a pool of plasmids (A), each with a human cDNA inserted into the 3′-UTR after a thymidine kinase-zeocin fusion protein (TKzeo; Fig 4). Cells are transfected with Target ID Library (B) and allowed to recover for 3-5 days. Constructs can integrate into the genome during this recovery period (C), and express the encoded transcript (D). After recovery, cells are exposed to zeocin (E). Cells expressing the TKzeo fusion protein from stably integrated Target ID constructs survive zeocin selection (F). Untransfected cells die (G). In addition, any cells containing a construct that is a target for an endogenous miRNA or any other factor expressed in the cell that inhibits expression of TKzeo from the Target ID construct will die (H).
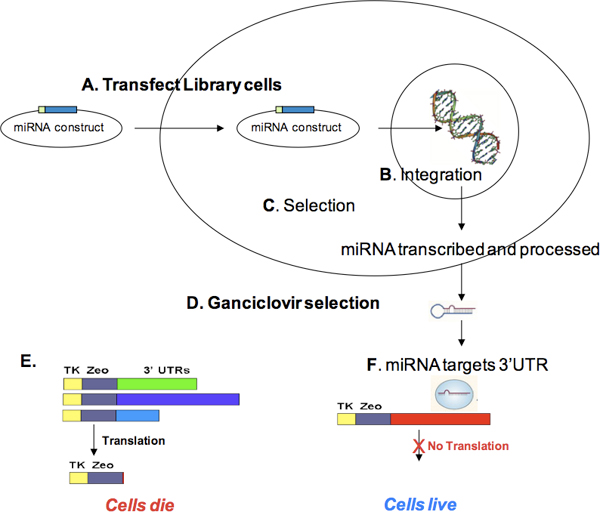
Figure 3. miRNA transfection and ganciclovir selection. Cells containing the Target ID Library (i.e., zeocin-selected cells) are transfected with a selectable microRNA expression construct (A). During recovery, the miRNA expression construct can integrate (B) and express the selectable marker encoded on the miRNA construct. After selection for stable integration (C) and cell expansion, cells are treated with ganciclovir (D). Cells producing thymidine kinase (TK) in the presence of ganciclovir (i.e., cells expressing TKzeo constructs not targeted by the miRNA) will die (E) On the other hand, cells containing Library constructs with miRNA target sites will not produce TK, and therefore, will survive ganciclovir selection (F). Surviving cells can be grown, gDNA isolated, and the cDNA containing miRNA target sites PCR-amplified using the Target ID Amplification primers. PCR products may be sequenced and aligned with the human genome to identify miRNA targets.
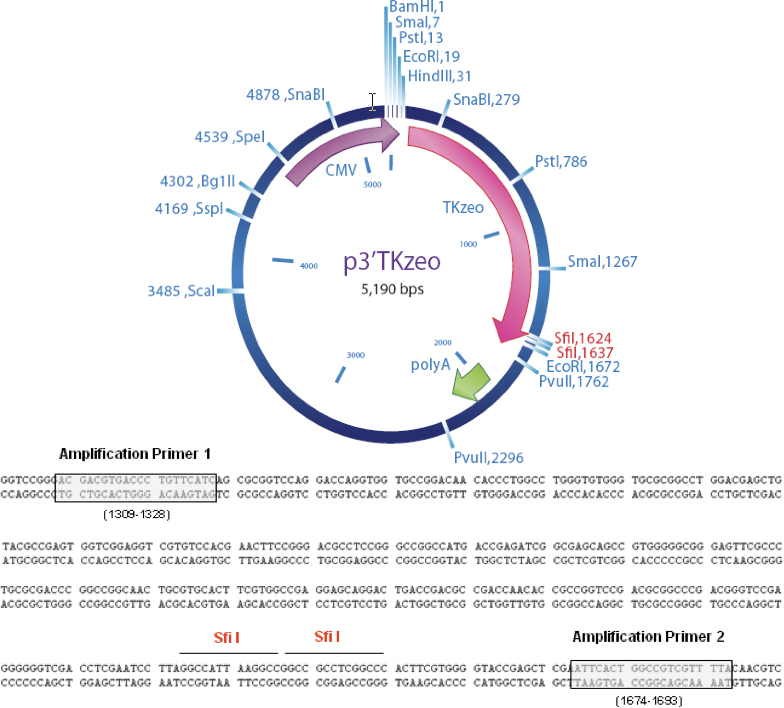
Figure 4. Plasmid Map and Location of Amplification Primers. Sfi I are the sites of cDNA Cloning.
Primer Sequences
MISSION Target ID Amplification Primer 1
5΄ ACGACGTGACCCTGTTCATC 3΄
MISSION Target ID Amplification Primer 2
5΄ TAAAACGACGGCCAGTGAAT 3΄
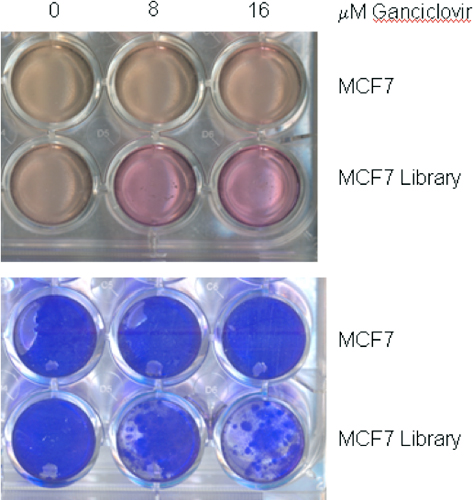
Figure 5. Ganciclovir effect on cell growth. Twenty-four well plates were seeded with two-hundred thousand MCF-7 cells or MCF-7 cells containing the Target ID Library. Twenty-four hours later, the medium was replaced with medium containing 0, 8, or 16 μM ganciclovir. After 15 days, the plate was photographed (6 upper wells), then the wells washed with HBSS and stained with Brilliant Blue R Staining Solution (B6529) (6 lower wells). Note that ganciclovir has no effect on MCF-7 cells without Library because they do not express thymidine kinase (TK). On the other hand, MCF-7 Library cells do express TK but are not completely killed by ganciclovir at 8 or 16 μM, as shown by the live cells taking up Brillinat Blue. However, the Library cells do stop growing in ganciclovir, as evidenced by color of the phenol red dye in their medium. The arrested cells do not acidify their medium, and the color remains reddish instead of changing to orange-yellow.
| Reagent | Volume / Reaction |
| Jumpstart REDTaq ReadyMix Reaction mix | 10 μl |
| Amplification Primer 1(25 μM) | 0.2 μl |
| Amplification Primer 2 (25 μM) | 0.2 μl |
| Genomic DNA | 50-200 ng |
| Water, Molecular Grade | to 20 μl |
Table 1.
| Step | Temp. | Time | Cycles |
| Initial Denaturation | 95 °C | 5 min. | 1 |
| Denaturation | 95 °C | 30 sec. | 40 cycles |
| Annealing* | 62 °C | 30 sec. | |
| Extension | 68 °C | 2 min. | |
| Final Extension | 68 °C | 5 min. | 1 |
| Hold | 4 °C | Hold |
Table 2.
*Annealing temperatures may vary, but we have observed the best amplification products with annealing temperatures ranging between 53 and 64 °C.
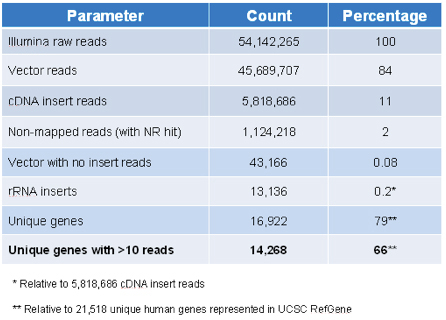
Table 3. Target ID Library content by Illumina sequencing.
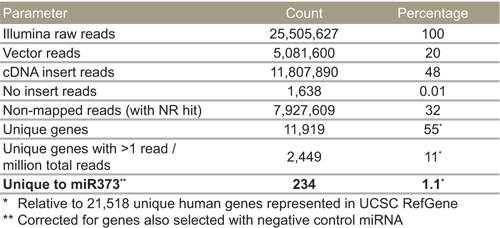
Table 4. miR-373 selected targets by Illumina sequencing.
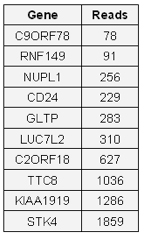
Table 5. 10 targets previously identified by RNA microarray.
