1. Coating of plates with ECM proteins
- Under sterile conditions, mix 120 μg of laminin with 12 ml of cold sterile 1X PBS per plate.
- Take Bioflex untreated plates and add 2ml of the solution to each well (final concentration of laminin is 2 μg/cm2). Alternatively, other ECM proteins could be used, such as collagen-1 [10 μg/cm2], fibronectin [5 μg/cm2], vitronectin [0.5 μg/cm2] or elastin [10 μg/cm2]. The coating technique is similar to what it is described for laminin-coating. Make sure the wells are completely covered by the solution.
- Wrap the plates with plastic and put at 4°C on a flat surface overnight to allow the laminin to adsorb to the bottom of the wells. In these conditions, plates can be stored for at least one week while still retaining good ECM function.
- Next day, under sterile conditions, wash the wells 3 times with 1X PBS.
- Add 1ml of 1% BSA in 1X PBS to each well and incubate at 37°C for 1h to block non-specific binding sites on the membranes.
- Rinse the plates 3 times with 1X PBS to remove unadsorbed proteins.
- Keep the plates at 37°C in DMEM until cells are isolated from step 2.14.
2. Isolation of fetal rodent lung type II epithelial cells
The day before isolation have the screen cups with different size nylon meshes autoclaved and ready.
- Obtain fetal lungs from timed-pregnant mouse/rat on E17-19 of gestation. Transfer the tissue into a 50 ml conical tube containing DMEM medium and place it on ice.
- Make digestion buffer in a 50 ml conical tube: (10 ml). For this volume, lung tissue from approximately 20 fetuses from mouse/rat is used.
8.5 ml DMEM
0.5 ml 500mM pH 7.4 HEPES
5 mg Collagenase 1
5 mg Collagenase 1A
1 ml Chicken serum, heat inactivated - Mix well until all components are dissolved and filter through a 0.2 micron syringe filter inside the sterile hood. Place the conical tube containing the digestion buffer in a water bath at 37°C.
- Remove the DMEM medium from the conical tube from step 2.1 and transfer the tissue into a sterile Petri dish.
- Mince the lungs well with sterile scissors or razor blade to make tissue pieces less than 1 mm and transfer the tissue into the conical tube containing the pre-warmed digestion buffer from step 2.3.
- Digest the tissue by pipetting up and down:
100 times with 10 ml pipette
100 times with 5 ml pipette
100 times with Pasteur pipette - Centrifuge the homogenate at 1,300 rpm for 5 minutes at room temperature.
- Carefully remove the supernatant by aspiration and re-suspend the pellet containing the cells in 15 ml of DMEM plus 20% FBS.
- Take out the screen cups with 100-, 30- and 15-micron nylon meshes and place them on sterile 150 ml beakers.
- Add cell homogenate from step 2.8 and sequentially filter through 100-, 30-, and 15-micron mesh screen cups.
- Collect clumped non-filtered cells from the 30- and 15-micron meshes after several washes with DMEM plus 10% FBS to facilitate the filtration of non-epithelial cells.
- Discard the filtrate from the 15-micron mesh which contains mostly fibroblasts.
- Purify further type II cells by incubating cells from step 2.11 in 75-cm2 flasks for 30 min (approximatelly10 ml of cell suspension per flask).
- Collect the supernatant and centrifuge at 1,300 rpm for 5 minutes at room temperature to pellet the cells.
- Carefully remove the supernatant by aspiration and re-suspend the pellet containing the cells in serum-free DMEM. The volume of resuspension depends on the amount of tissue processed. Approximately, we plate the equivalent of cells obtained from one fetus in each well (2 ml) in order to achieve between 60-70% confluency the following day.
- Plate the cell suspension on Bioflex six-well plates precoated with laminin or fibronectin.
3. Isolation of fetal rodent lung fibroblasts
- Follow the same steps described above for type II cells isolation up to 2.11.
- Take the filtrate from the 15-micron mesh (without previous washing; approximate volume is 10-15 ml) and plate onto 75-cm2 at 37°C for 30-60 min to allow fibroblasts to adhere.
- Aspirate the supernantant and replace with serum-free DMEM and incubate overnight.
- Next day, harvest the cells with 0.25% (wt/vol) trypsin in 0.4 mM EDTA and plate them on Bioflex plates precoated with fibronectin.
4. Experimental system to provide mechanical stimulation to lung cells
- Seed the cells (around 50% confluent) on Bioflex culture plates in serum-free DMEM (2 ml/well) and let them attach and spread for at least 6h before experiments. In general our laboratory maintains cells in culture for around 24h before applying mechanical strain. If a specific number of cells are required for the experiments, after isolation, type II cells are maintained in serum-free DMEM 75-cm2 flasks and the next day, cells are trypsinized, counted and then seeded.
- Next day, media are replaced with fresh serum-free DMEM (2 ml/well) and bioflex plates are mounted in a Flexcell FX-5000 Strain Unit. Monolayers should be no greater than 80% confluent before the initiation of the experiments.
- Equibiaxial strain is applied to the membranes. The regimen of strain varies depending on simulation of mechanical forces in vivo (see discussion).
- Cells grown on non-stretched membranes cultured in parallel are used as controls.
- At the end of the experiments, monolayers can be processed to analyze changes on gene expression (for example surfactant protein C) by real time-PCR, protein abundance by Western blot, etc. Also, monolayers can be fixed for immunocytochemistry experiments. For this technique, after fixation, silastic membranes are cut out from the plates and mounted on glass slides using 10-20 μl of water as a mounting agent before permeabilization and incubation with antibodies. Supernatant can also be used to investigate the presence of growth factors, cytokines, etc.
5. Representative Results
Figure 1 and Figure 2 show representative phase-contrast photographs of E18 fetal mouse type II cells isolated using the technique described in this manuscript.
Figure 3 demonstrates that mechanical strain induces differentiation of fetal type II epithelial cells using surfactant protein-C as a marker.

Figure 1. Representative phase-contrast photograph taken right after isolation showing the clumped appearance of fetal type II cells.
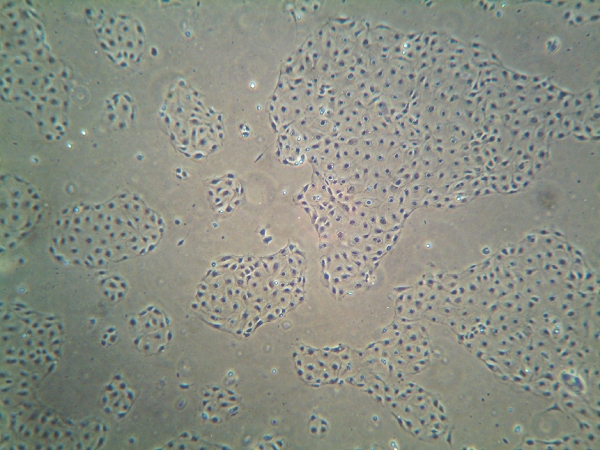
Figure 2. E18 fetal type II epithelial cells were isolated as described here and plated on bioflex plates coated with laminin. Photograph was taken the following day after non-stretched cells were fixed in paraformaldehyde. The purity of the cells was determined to be 90 ± 5% by microscopic analysis of epithelial cell morphology and immunostaining for SP-C.

Figure 3. Fetal type II epithelial cells were exposed to 5% cyclic strain at 40 cycles/min for 16h. A) Northern blot of surfactant protein C (SP-C) mRNA expression showing that strain induces type II cell differentiation using different ECM substrates. +/- signs represent exposure or not to strain, respectively. Data are presented as mean +/- SEM, n=3; * P<0.05. B) Fluorescence immunocytochemistry images demonstrating SP-C protein levels (green) in fetal type II cells exposed or not (control) to mechanical strain . Nuclei were counterstained with dapi (blue). Bar, 10 μm. C) Western blot results from three experiments showing that mechanical stretch increases SP-C protein. N=3; * P<0.05.
