1. Initial Characterization and Considerations
- Biofilm formation is generally influenced by cell density and growth rate. Therefore, it is necessary to determine the growth rate (increase in optical density (OD) over time) and yield (final cell number) of the strain(s) of interest by performing simple growth curve and cell plating assays. Defects in growth, or a lack of correlation between OD and cell number, must be taken into consideration when interpreting results from spotting experiments.
- Include the appropriate positive and negative controls on the same plate when assessing wrinkled colony formation, as minor plate-to-plate variation may impact biofilm development.
- Identify the best conditions for spotting, i.e., those that reveal the most distinct differences between the control and mutant(s) of interest. Grow strains in liquid culture under various conditions, such as different media or temperatures, and to different stages of growth (exponential or stationary phase) prior to spotting. Spot 10 μl of culture at various cell densities onto the appropriate media, and incubate at the desired temperature until colony morphologies become apparent.
- The end-point assay involves assessment of wrinkled colony formation at a predetermined time point after spotting (i.e., 48 hours). This assessment is useful for strains that exhibit (or are proposed to exhibit) severe defects in biofilm formation.
- The time course assay assesses wrinkled colony formation over a period of time (i.e., every hour) post-spotting. This assay permits a semi-quantitative assessment of biofilm formation by allowing the determination of the start of wrinkled colony formation and pattern development over a period of time. The duration of the experiment and number of collection points for a given time course should be determined in preliminary experiments.
- To easily keep track of the time post-spotting, set a timer to count up.
2. Microscopic Assessment of Wrinkled Colony Morphology in V. fischeri
- Inoculate V. fischeri cells into 5 ml of LB-Salt (LBS) medium 13 (1% [w/v] tryptone, 0.5% [w/v] yeast extract, 2% [w/v] sodium chloride, 50 mM Tris-HCl [pH 7.5]) containing any necessary antibiotics and incubate, with shaking, overnight at 28 °C. In the morning, subculture the cells with a 1:100 dilution into 5 ml of fresh LBS medium and incubate under the same conditions until the cells have reached the desired OD600 (e.g., OD600=0.2 or 0.5).
- Pipette 1 ml of culture into a microfuge tube and centrifuge in a microfuge set at maximum speed for one minute. Remove the supernatant by aspiration. Wash the cells to remove residual media and extracellular components by resuspending the pellet in 1 ml of sterile 70% artificial seawater (ASW) (35 mM MgSO4-7H2O, 7 mM CaCl2-2H2O, 210 mM NaCl, 7 mM KCl) and repeating the centrifugation. Resuspend the washed pellet in 1 ml of 70% ASW.
- Make sure that each sample contains the same number of cells, as estimated by the OD at 600 nm. Make any necessary adjustments by diluting more concentrated samples with additional 70% ASW. In preliminary experiments, spot cultures at various initial OD values to determine an optimal starting OD for a given set of strains or conditions. The best results are obtained for V. fischeri when spots are generated from cultures with an OD of approximately 0.2.
- Spot 10 μl of the washed cells onto LBS plates containing any necessary antibiotics. When spotting a culture onto a plate, make sure to steady the pipette (with your finger) just above the agar surface. Spot vertically, not at an angle, and eject the liquid slowly for uniform distribution of the spot. Typically each strain is spotted once per plate (up to 6 spots per plate), with multiple plates (2-3) per experiment.
- To ensure that the spotted culture remains evenly distributed, allow the spot to dry before moving the plate to the incubator. Invert the plates and incubate them at 28 °C.
- Monitor the morphology of the growing spot hourly beginning at 12-15 hours following inoculation. We use a dissecting scope (Zeiss stemi 2000-C) with a camera attachment (ProgRES C10PLUS) and the ProgRes CapturePro and ImageJ software programs to observe and document colony morphology and to assess the start and progression of wrinkled colony formation.
- For the specific setup listed in step 2.5, the spots are illuminated from underneath through a transparent glass stage with a CL 1500 EC cold light source, while the images are captured from above. This setup is optimal for imaging wrinkled colony development because V. fischeri colonies (spots) are translucent.
- In the absence of the specific equipment listed in step 2.5, monitor wrinkled colony morphology using any dissecting microscope that allows for full colony viewing and has an adjustable light source and, ideally, an attached camera. If necessary, a digital camera can also be utilized in the absence of an attached camera, but this is not optimal.
- To best visualize wrinkled colony development, it is necessary to adjust both the lighting intensity and angle of reflection underneath the bacterial colonies such that the three-dimensional morphology of the developing biofilms can be discerned. Determine the optimal lighting conditions that provide the strongest contrast between the spotted colony and the surrounding agar background, such that the architecture (wrinkles) of the colony is clearly distinguishable.
- In some cases, the color of the spotted colony must also be taken into consideration, as alterations in colony color can occur during biofilm formation. Adjust the intensity and angle of the lighting source to reveal these subtle changes in coloration. Once the appropriate settings are achieved, maintain them for the duration of the experiment.
- Note how much time elapses from the time of inoculation to the time at which wrinkled colony formation initiates for each strain or condition. Define the start of wrinkled colony formation as the time point at which the formation of patterning and 3D structures (i.e., striations forming at the outer edge or ‘ripples’ occurring in the center) is first apparent. Mutant cells may exhibit decreased or increased time to the start of wrinkled colony formation (e.g., Fig. 2).
- Document the start and development of wrinkled colony formation by capturing appropriate digital images. It is important to use the same magnification when collecting images throughout the experiment. Switch the view from the eyepiece of the microscope to the computer screen using the lever located on the back of the camera. When switching between eyepiece and computer screen viewing, adjust the view and focus accordingly.
- Before imaging spotted cultures, the lid of the Petri plate is typically removed to provide the clearest image. However, this is an optional step, as spotted cultures can be imaged through the lid using the setup outlined in steps 2.5 and 2.6.
- At each time point following the start of wrinkled colony formation, note the pattern of wrinkled colony development. The architecture may develop from the inside out, or the outside in. This assessment provides a mechanism to distinguish the biofilms formed by different strains or under different conditions.
- Measure the diameter of the developing colony at each time point. This can be done manually, or digitally using an associated software program. If done digitally, use the ProgRes CapturePro software program and first calibrate the scale bar to the specific magnification used in the experiment. Include the scale bar in each image captured.
- To calculate the colony diameter using the ImageJ software program, open each image file. Standardize the scale bar as follows: Select the “Straight line option” from the toolbar. Overlay the embedded scale bar with a straight line of the same length and width. Select “Set Scale” from the tab labeled “Analyze.” In the “known distance” box, insert the corresponding length (as determined from the original scale bar; i.e., 2 mm) and select “OK”.
- Use the “Straight line” option to insert a horizontal line across the spot. Under the Analyze tab, select “Measure”. In the new Results window, the calculated length will be provided. Obtain a second measurement by inserting a perpendicular line across the spot. Re-calculate the diameter. Record the average of the two measurements. Graph the average diameter for each spot at each time point using a software program such as Excel.
- The end of the experiment occurs either at a specified time point or when there is no further development of the biofilm. Once the end is reached, consolidate the images into a figure(s) to visualize the pattern development over time using a software program such as Powerpoint.
3. Representative Results
In these experiments we used V. fischeri as a model organism to study biofilm formation by evaluating the development of wrinkled colonies on a solid agar surface. Biofilm-producing strains of V. fischeri form colonies with extensive 3D architecture within 40 h (Fig. 1) 8,9,14. When examined over a time course, it becomes apparent that wrinkled colony formation by the control strain initiates as early as about 12 h post-inoculation (depending on the specific conditions) (Fig. 2) 9. In contrast, biofilm formation by a representative mutant is delayed by approximately 4 h, not initiating until approximately 16 h post-inoculation 9. Repeats of these experiments suggested that the timing was relatively consistent, making this assessment semi-quantitative 9. A second semi-quantitative measure of biofilm formation makes use of the change in diameter of the developing colony over time. As shown in Fig. 3A, a representative biofilm-competent strain forms colonies that increase in complexity and diameter relative to a representative strain that does not form biofilms. Careful measurements over time of the diameters of the colonies formed by these two strains revealed that the size of the biofilm-competent colonies increased at a greater rate than the biofilm-negative colonies, and at the end time point the two differed by almost 2-fold (Fig 3B). Thus, although images of a representative late time or “end-point” colony morphology frequently are shown in the literature, additional, semi-quantitative data can be collected that will permit a better understanding of the biofilm defect.
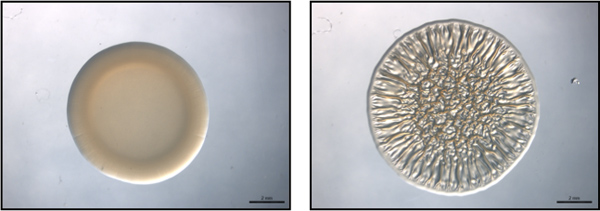
Figure 1. End-point assay. This figure is an example of an end-point assay using representative non-biofilm-forming (left) and biofilm-forming (right) strains of V. fischeri. These images were collected at 40 h post-spotting. These images were generated with wild-type V. fischeri containing a vector control (non-biofilm-forming) or an rscS overexpressing plasmid (biofilm-forming) and were part of the data set collected for Morris et al., 2011.
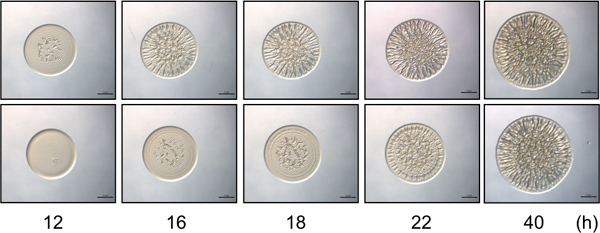
Figure 2. Time-course assay. The upper panel contains representative images of biofilm formation by a biofilm-competent control strain of V. fischeri over a selected time course. Initiation of biofilm formation is evident at 12 h. The lower panel contains representative images of a mutant strain of V. fischeri that exhibits a delay (4 h) in the start of wrinkled colony formation over time, with biofilm formation initiating at 16 h post-inoculation. Note that at 40 hours the strains look similar in the intensity and patterning of biofilm formation, while subtle differences between these strains are only observed at the earlier time points. These images were generated with rscS overexpressing wild-type and sypE mutant V. fischeri cells and were part of the data set collected for Morris et al., 2011.
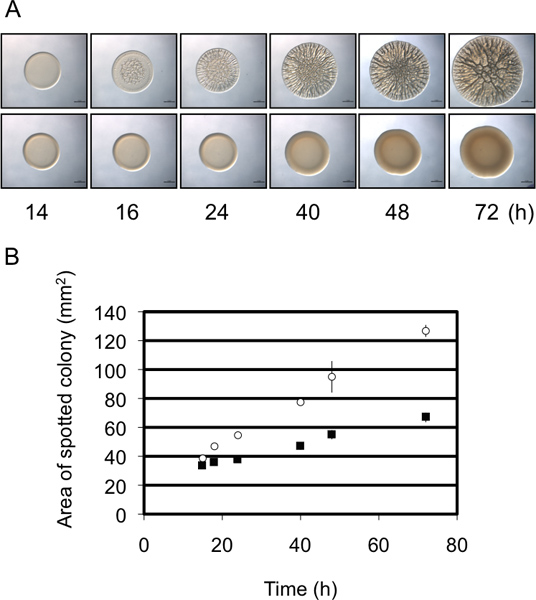
Figure 3. Colony diameter as a semi-quantitative analysis of biofilm formation. (A) Time course of biofilm formation by representative biofilm-forming (upper panel) and non-biofilm-forming (lower panel) strains of V. fischeri. Note that the biofilm-forming strain exhibits a greater increase in diameter over time relative to the non-biofilm forming strain. These images were generated with wild-type V. fischeri containing an rscS overexpressing plasmid (biofilm-forming) or a vector control (non-biofilm-forming) and were part of the data set collected for Morris et al., 2011. (B) A graphical representation of the increase in colony diameter over time by the two strains in panel A. These data were generated using the ImageJ and Excel software as outlined in the protocol.
