The weights of animals that were used in this study ranged from 100-200 g. Following the described protocols, we conducted electrophysiological measurements and molecular analysis of neurons of abdominal ganglia isolated from animals ranging from 2-5 g to 200-300 g.
Standardization of protease treatment is important for successful electrophysiological measurements of neurons in the ganglia. Initially, multiple protease (Dispase) concentrations and durations were used and bursting action potentials from R15 neuron were recorded26,31. These measurements were compared to direct (without protease treatment) recording from the intact ganglia. Concentration of the protease (0.1% protease, see step 1.3) that produced bursting potentials that were comparable to those obtained from exposed ganglion without protease were used in the subsequent experiments.
Using the protocol described, action potentials (AP) in response to Ach exposure to R15, a cholinergic neuron and L7, a motor neuron that does not respond to Ach were recorded. As expected, simultaneous electrophysiological recordings show that Ach induce depolarization in R15 whereas there was no change in firing of L7. Figure 2 shows the application of Acetylcholine (Ach) directly into the cell chamber. 1 M Ach (volume 0.3 μl) was directly applied into the chamber using micropipette in order to obtain a final 1 mM concentration inside the chamber.
Ach was washed off from the bath by perfusion and recorded again from R15 neurons. Figure 3 shows waveforms suggesting that Ach effect is reversible. Analysis of the parameters AP in control, during depolarization and after wash is shown. During depolarization amplitude is decreased and duration of AP increased significantly.
Following electrophysiological measurements, R15 and L7 neurons were isolated and RNAs were prepared for qPCR analyses. A linear T7 RNA Polymerase—driven transcription using MessageAmp II aRNA Amplification Kit from Ambion was used for amplifying RNAs from single neurons. For comparison, identified neurons R2 and L11 and the whole abdominal ganglia from adult (6-9 months old) and young animals (1-2 months old) were also studied. qPCR amplification were carried out in a 7900HT Fast Real-Time PCR System under the following conditions: 95 °C for 10 min, followed by 40 cycles of 95 °C for 15 sec, 60 °C for 1 min. There were four biological replicates in the experiment. Each biological replicate had four technical replicates in the qPCR set up. Ct values from four replicates were averaged and 1/Ct was calculated to find the absolute expression level of CREB1 in each sample (Figure 4). An estimation of the absolute expression levels of CREB1 gene in young and adult Aplysia ganglia, and in four identified single neurons was monitored by the Ct value obtained from qPCR measurements.
The starting mRNA concentration was the same in both young and adult abdominal ganglia. The same forward and reverse CREB1 primer set were used in all qPCRs, both in studies using ganglia and single neurons. The Ct values are very similar between young and adult abdominal ganglia. Similarly, Ct values were measured in the case of single neurons where the starting total mRNA concentrations were low and two rounds of amplification yielded a good amount of RNAs for cDNA synthesis. The Ct values of CREB1 amplification suggested that CREB1 expression in R15 and L7 neurons are comparable whereas R2 and L11 showed significant differences in expression when compared to L7 or R15 (Student's T test, p<0.05) (Figure 4).
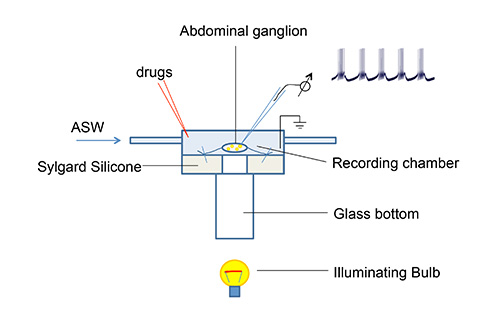
Figure 1. Microscope stage and perfusion system for ganglia electrophysiology. Perfusion system and stage were made using Plexiglas and are transparent. This setup is mounted on to a metal block (not shown) to hold position. The metal block is designed such that samples can be illuminated from the bottom. A sample electrophysiological bursting pattern recorded from R15 neuron is shown.
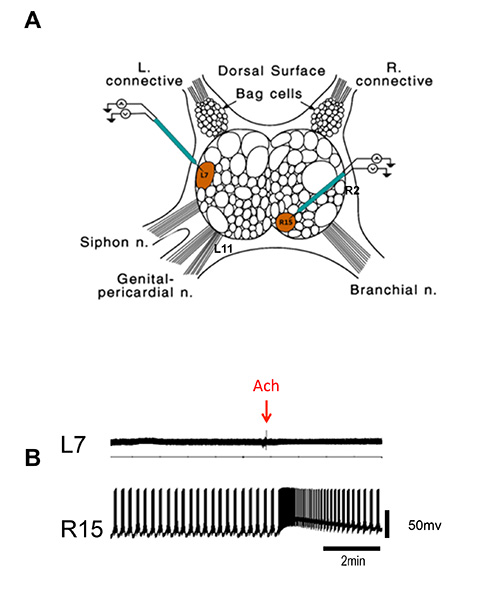
Figure 2. Simultaneous recording from L7 and R15 neuron of abdominal ganglia. (A) Cartoon of abdominal ganglia showing the position of L7 and R15 neurons (modified from Kandel 2001, reprinted with permission from AAAS). Also shown are the positions of R2 and L11 in the abdominal ganglia. (B) Representative intracellular recordings from L7 and R15. Arrow indicates time of Ach application. One could continue these measurements for 8-10 hr.
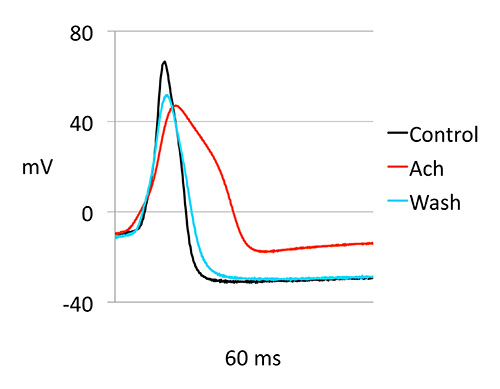
Figure 3. Ach induced changes in action potential (AP) of R15. AP waveforms were analyzed before, during and after Ach exposure. Representative waveforms are shown. ms: millisecond, mV: millivolt.
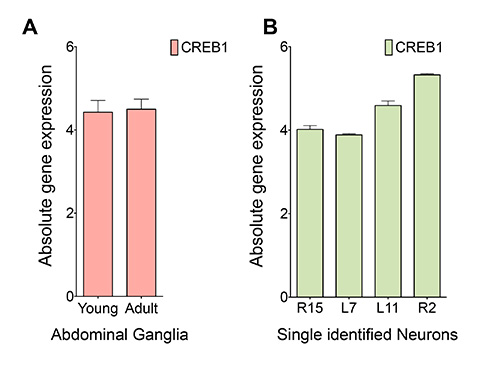
Figure 4. qPCR analysis of expression of CREB1 in abdominal ganglia and single identified neurons. (A) Absolute quantification of CREB1 transcript in the young and adult abdominal ganglia of Aplysia. Total RNA was isolated using standard Trizol protocol as described in the methods. 1 μg of total RNA was used to synthesize cDNA, followed by real time PCR amplification using CREB1 primers. (B) Absolute quantification of CREB1 transcript in identified single neurons of old Aplysia. Total RNA was subjected to two rounds of in vitro amplification before cDNA synthesis. 1 μg of aRNA was used to synthesize cDNA, followed by real time PCR amplification using CREB1 primers.
