When age-matched, genetically identical C. elegans are exposed to P. aeruginosa, a wide distribution is observed in levels of colonization (Figure 1A). With the help of the protocol described here efficient separation of noncolonized from colonized worms can be achieved (Figure 1B). Unlike noncolonized worms, colonized worms show signs of damage, such as sluggishness and reduced defecation19. The latter may be a reason why worms colonized by P. aeruginosa have difficulty clearing infection. This has direct implications for the described protocol suggesting that worms sorted as noncolonized were not previously colonized. To directly test whether this assertion is correct, hand-picked colonized worms were subjected to similar washing steps as those in the sorting experiment and subsequently examined under a fluorescent microscope; only 4/56 cleared the pathogen. Therefore, it is likely that the proportion of previously-colonized worms among those sorted as noncolonized is negligible. However, it is advisable to monitor changes in colonization during washing steps to prevent effects of mistaken sorting on analysis, which may be more of an issue for other pathogens than P. aeruginosa.
Previous work employing the protocol described here used sorted worms for a microarray gene expression analysis and observed essentially identical responses to P. aeruginosa in colonized and noncolonized worm populations that had been exposed to the pathogen for the same amount of time19. Figure 2 demonstrates the same trend using quantitative (q)RT-PCR to measure expression of four genes, known to be part of the C. elegans immune response: lys-2 encodes a lysozyme, F08G5.6 and F55G11.2 are two uncharacterized genes, that respond specifically to infection, of which the first also contributes to infection resistance, and pgp-5, which responds to infection as well as to heavy metal stress. All four genes were induced similarly by P. aeruginosa exposure, regardless of the colonization status of the worms. This corroborates previously published results indicating that colonization and its associated damage are not required for an immune response, instead pointing at pathogen-associated molecular patterns as the signal. That pgp-5 is induced in noncolonized animals, which are not expected to be experiencing extensive damage, suggests an overlap in the regulatory programs controlling immune responses and general stress responses in C. elegans.
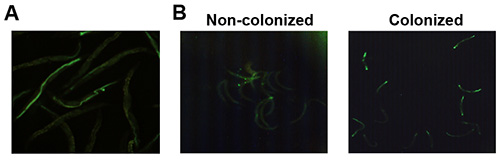
Figure 1. Colonized and noncolonized C. elegans separated using a wormsorter. (A) Representative image of wild-type adult C. elegans exposed to GFP-expressing P. aeruginosa for 18 hr. (B) Representative images of a noncolonized and colonized worms separated using the wormsorter. Please click here to view a larger version of this figure.
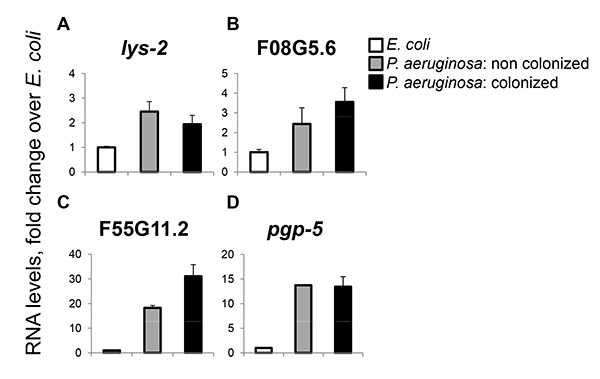
Figure 2. Colonization is not required for the transcriptional response to P. aeruginosa. RNA levels of lys-2 (A), F08G5.6 (B), F55G11.2 (C), and pgp-5 (D) measured by qRT-PCR in wild-type animals exposed to E. coli or P. aeruginosa for ~18 hr. Shown are means and SD of duplicate measurements from one experiment, which is a representative of three. Please click here to view a larger version of this figure.

Table 1. Materials and reagents needed for a wormsorter experiment.
