As illustrated in Figure 1, the mini-himar1 mariner transposon vector, pFAC, contains two 27 bp inverted repeats with TA insertion sites flanking the aacC1 gentamycin resistance cassette, with its σ70-dependent promoter, and a multiple cloning site (MCS). Additionally, the pFAC vector contains genes encoding the highly active himar1 transposase, β-lactamase (bla), and the tra transfer operon. The TA insertion sites allow for an efficient integration into the genome of P. aeruginosa strains. Accuracy in identifying and selecting mucoid mutants is critical for the overall success in determining gene loci involved with alginate regulation. Examples of mucoid and nonmucoid isolates are shown in Figure 1.
To more accurately identify mucoid mutants, the completed conjugations should be plated on multiple large plates (150 mm). The cell mixtures should be resuspended in sterile PBS and plated by spreading onto the PIA plates plus gentamycin (300 µg/ml). Dilutions of cell mixture in PBS will be spread on the plates with the goal of having between 1,000-3,000 colonies/plate. Based on the protocol above, each conjugation should produce a bank of ~20,000 mutants over 15-20 large plates. Optimal incubation time for the plates at 37 °C is between 24-36 hr. Due to the overproduction of alginate, the mucoid colony should be clear or white in color and have creamy or slimy appearance. If mucoid colonies cannot be discerned, then incubate for an additional 24-36 hr at room temperature. The mucoid mutants are best isolated by passing three times on a regular PIA plate with gentamycin. During this time, you can determine the stability of the mucoid phenotype. The purified mucoid colony will be used for genomic DNA extraction and identification of the genes by iPCR. iPCR products can be visualized using agarose gel electrophoresis (Figure 2). If successful, there should be a single amplicon at the appropriate size. PAO1 genomic DNA that has undergone restriction digestion and ligation can be used as a negative control. As illustrated in Figure 2, there is an absence of iPCR amplicon in PAO1, however we detect a single amplicon of ~1,400 bp and ~1,000 bp in PAO1-VE2 and PAO1-VE13, respectively. The size of these amplicons corresponds to the insertion of the himar1 mariner transposon into the promoter region of PA4033 (mucE) in PAO1-VE2 and internally in PA5484 (kinB) in PAO1-VE13. DNA sequencing of the iPCR amplicons should be carried out with the GM5OUT primer. When using the Basic Local Alignment Search Tool (BLAST) to analyze the target sequence, the inverted repeat on the 5' end of pFAC and TA dinucleotide mark the site and orientation of the himar1 insertion in the genome of P. aeruginosa (Figure 2).
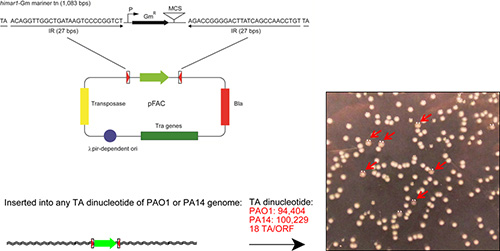
Figure 1. Schematic diagram of mini-himar1 mariner transposon vector, pFAC, and a representation of mucoid mutants. Plasmid pFAC contains a himar1 mariner transposon element with two inverted repeats, and a gentamycin resistance cassette (aacC1) for selection, a gene encoding the hyperactive himar1 transposase, and a conditional replicon. The mini-himar1 mariner transposon can cause high-density insertion in P. aeruginosa because of the abundance of substrate (TA dinucleotide). The number of TA dinucleotides in the genomes of two P. aeruginosa strains, PAO1 and PA14 are shown in red. The red arrows indicate the mucoid mutants identified using this procedure. Please click here to view a larger version of this figure.
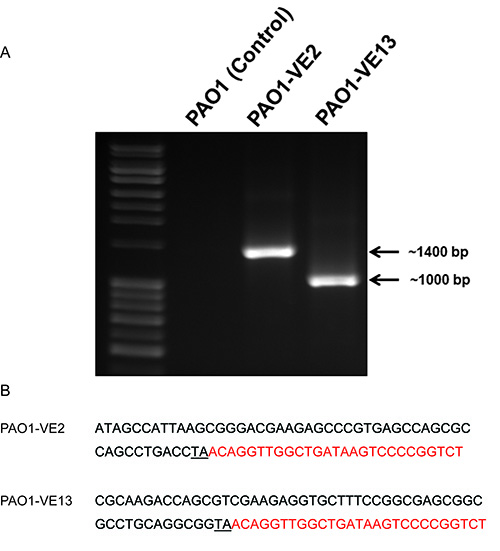
Figure 2. Representative iPCR and sequencing results. A) 1% agarose gel electrophoresis of iPCR amplification using the Forward Gm3OUT and Reverse Gm5OUT primers and a 1 kB ladder: PAO1 (negative control), PAO1-VE2 (1396 bp), and PAO1-VE13 (999 bp). The genomic DNA from three strains of P. aeruginosa was extracted and digested with SalI restriction enzymes followed by self-ligation. The closed circular DNA was used as templates for iPCR as described in the text. B) Comparative sequence analysis using the PAO1 reference genome determines the precise location of the himar1 mariner transposon insertion in PAO1-VE2 and PAO1-VE13. The sequence labeled in red indicates the 5' end of inverted repeat in pFAC. The insertion site of TA was underlined. A BLAST search of these two sequences will map the exact location and orientation of the himar1 transposon within the genome of PAO1.
