The method that is described above reliably produces high quality material for analysis with in situ probes, transcriptional reporters, protein traps and antibodies. In Figure 1 we display eye-antenna, genital, wing, haltere and leg discs that are routinely recovered with this method. These discs have been treated with a phalloidin-conjugated fluorophore, which binds to F-actin and therefore outlines each cell. If the tissue has been fixed properly then the morphogenetic furrow of the eye disc, the edges of the concentric tissue folds in the genital, antennal and leg discs, and the dorsal-ventral axis of the wing and haltere discs will all appear as sharp edges. If a tissue is properly fixed then antibodies, fluorescent proteins and in situ probes will also reveal sharp patterns. Several examples are shown in Figure 2. The top three panels display discs that have been stained with different antibodies while the lower three panels show discs in which GFP is used to mark populations of cells.
One of the most striking features of the eye-antennal disc is the morphogenetic furrow (Figure 1A), which can be seen as an indentation within tissue running along the dorsal-ventral axis 1, 22. Prior to the third larval instar all cells within the developing eye are unpatterned, undifferentiated, and morphologically indistinguishable from one another. At the start of the third larval instar the morphogenetic furrow initiates at the posterior margin of the eye field and progresses anteriorly towards the eye/antennal border 22. As the furrow progresses across the eye field the sea of disordered cells is transformed into an ordered array of periodically spaced unit eyes or ommatidia (Figure 1A) 22-23. Ahead of the furrow a gene regulatory network that includes the Pax6 homolog Eyeless (Ey) channels cells towards an eye fate (Figure 2A) 15. The initiation and progression of the furrow itself is dependent upon the activities of the Hedgehog (Hh) and Decapentaplegic (Dpp) signaling pathways 24-29. Indeed a dpp-lacZ reporter faithfully reflects the expression of the dpp locus within the furrow (Figure 2B) 30-31. As cells exit the furrow and begin to adopt their terminal fates, they express cell specific markers such as embryonic lethal abnormal vision (elav), which encodes a pan-neuronal RNA binding protein (Figure 2C) 32-34.
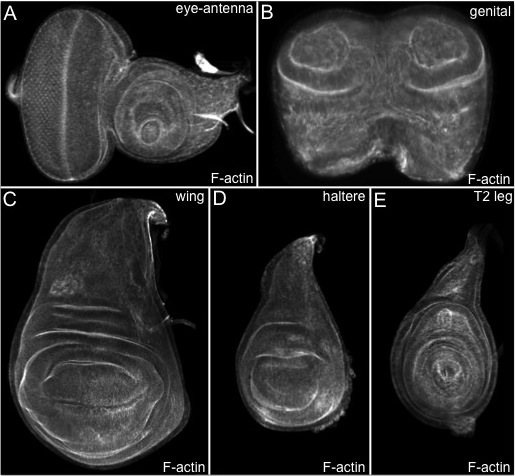
Figure 1. The imaginal discs of Drosophila melanogaster. (A–E) Confocal images of wild type eye-antenna, genital, T2 leg, wing and haltere imaginal discs. (A) As the morphogenetic furrow progresses across the eye field, a sea of unpatterned and undifferentiated cells is transformed into columns of unit eyes that are also called ommatidia. All discs are treated with phalloidin-conjugated fluorophores, which bind to and reveal F-actin distribution. Anterior is to the right and dorsal is up.
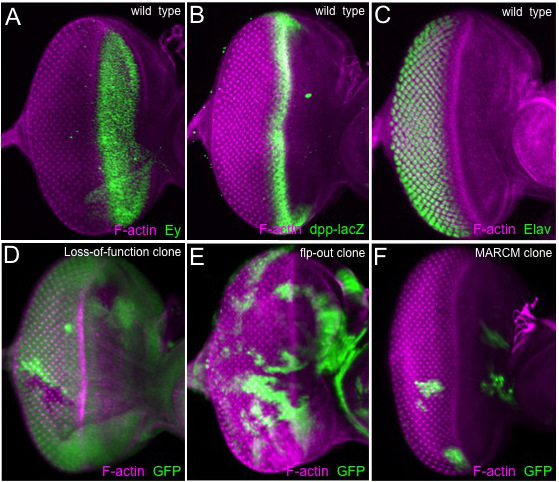
Figure 2. Clonal and expression analysis of the eye imaginal disc. (A–F) Confocal images of eye imaginal discs. (A) The Pax6 protein Eyeless (Ey) is distributed broadly ahead of the morphogenetic furrow. (B) A dpp-lacZ transcriptional reporter responds to Hh signaling and is expressed within the morphogenetic furrow. (C) The pan-neuronal RNA binding protein Elav is distributed in all developing photoreceptors behind the morphogenetic furrow. (D) An eye disc containing loss-of-function clones generated by the FLP/FRT system. The clones are identified by the lack of GFP (E). An eye disc containing over-expression clones generated with the flp-out system. Clones are positively marked with GFP. (F) An eye disc containing MARCM clones. Like the flp-out system, the MARCM clones can be identified by the presence of GFP. All detected proteins and genotypes are listed within the figure. Anterior is to the right and dorsal is up.
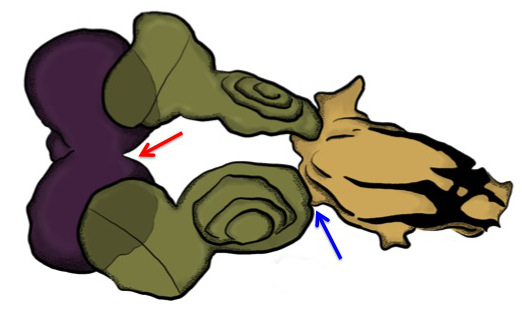
Figure 3. Eye-antenna-brain complex. A schematic drawing of the first day coarse dissection products. The only tissues that should be fixed are the mouth hooks, eye-antenna discs and brain (often times the ventral ganglion will remain attached as well — not shown). Purple = brain, green = eye-antenna discs, brown = mouth hooks. Anterior is to the right. If dissecting leg, wing, haltere and genital discs, the outer cuticle of the larva should still be attached to these tissues. Do not remove the overlying cuticle as you might lose the imaginal discs during subsequent transfers through various antibody, block and washing solutions. The excess tissue can be removed during the fine dissection steps (4.1–4.2).
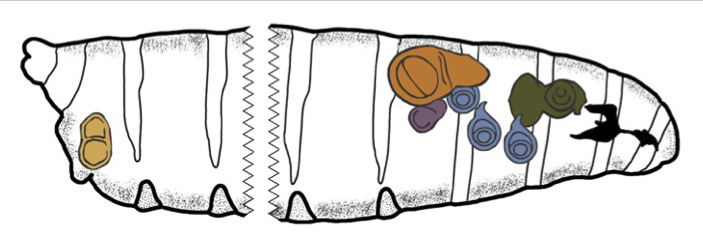
Figure 4. Position of imaginal discs within the Drosophila larva. A schematic drawing of the relative position of the eye-antenna, leg, wing, haltere and genital discs within a third instar larva. The eye-antennal disc is colored in green, the leg discs are in blue, the halter disc is in purple, the wing disc is in brown/orange and the genital disc is in light brown. Anterior is to the right.
| Name of Solution |
| 8% Paraformaldehyde |
| 0.2 M Sodium Phosphate Monobasic |
| 0.2 M Sodium Phosphate Disbasic |
| 1 N Sodium Hydroxide |
| 10% Triton |
| Distilled Water |
| 0.1 M Sodium Phosphate Buffer (Dissection Buffer) |
| 0.1 M Sodium Phosphate Buffer + 0.1% Triton (Wash Buffer) |
| Lysine Buffer |
| 2% Paraformaldehyde-Lysine-Periodate Fixative (PLP) |
| 10% Normal Goat Serum |
Table 1: List of Required Solutions.
| 8% Paraformaldehyde stock solution |
| To a 50 ml Erlenmeyer flask add the following: |
| 2.0 g paraformaldehyde |
| 23.0 ml of distilled water |
| 4.0 drops of 1 N sodium hydroxide (from a glass pasteur pipet) |
| Mix and heat on a stir plate until solution reaches a gentle boil |
| Allow to gently boil until paraformaldehyde is completely dissolved |
| Place on ice until cold (make fresh prior to each dissection) |
| 0.1 M Phosphate Buffer (Dissection (P) Buffer) |
| To a 50 ml conical tube add the following: |
| 18.0 ml 0.2 M Sodium Phosphate dibasic |
| 7.0 ml 0.2 M Sodium Phosphate monobasic |
| 25.0 ml distilled water |
| Store at 4 °C (1 week shelf life) |
| 0.1 M Phosphate + Detergent Buffer (Wash (W) Buffer) |
| To a 50 ml conical tube add the following: |
| 18.0 ml 0.2 M Sodium Phosphate dibasic |
| 7.0 ml 0.2 M Sodium Phosphate monobasic |
| 25.0 ml distilled water |
| 0.5 ml 10% Triton |
| Store at RT (1 week shelf life) |
| Lysine (L) Buffer |
| To a 50 ml conical tube add the following: |
| 0.4 g Lysine |
| 1.2 ml 0.2 M Sodium Phosphate dibasic |
| 8.0 ml Dissection (P) Buffer |
| 10.0 ml distilled water |
| Shake solution until lysine is completely dissolved |
| Place on ice until cold (make fresh prior to each dissection) |
| 2% Paraformaldehyde – Lysine – Periodate (PLP) fixative |
| 0.1 g Sodium Periodate |
| 15.0 ml of Lysine (L) buffer |
| 5.0 ml of 8% Paraformaldehyde |
| Shake solution well until sodium periodate is completely dissolved |
| Place on ice until cold (make fresh prior to each dissection) |
Table 2: Recipes for Required Solutions.
