1. Lipid Preparation
- Aliquot the desired amount lipids into a clear glass tube. Use E. coli polar lipid extract, but most lipid compositions can be used. If the lipids are in powder form, resuspend them in chloroform to a concentration of 20 mg/ml. Dry the lipids at RT under N2 gas until all solvent has evaporated.
- Resuspend the lipids in pentane and dry it again a steady stream of gas N2 to remove leftover traces of chloroform. Dry until all solvent has evaporated. While drying, gently rotate the tube so that the lipid is distributed in a uniform film covering the bottom third of the tube rather than a forming a dense mass at the bottom.
- Add the reconstitution buffer containing detergent (300 mM KCl, 50 mM citric acid, 25 mM K2HPO4, pH 4.5, 35 mM CHAPS) to dissolve the lipids. Use other mild detergents for this step if the protein is poorly tolerant to CHAPS.
- Resuspend the lipids to clarity using a water bath sonicator. Immerse the tube in the center of the water sonicator bath at maximum power in short (30 sec-1 min) pulses with short (30 sec-1 min) pauses. The solution should become clear.
NOTE: The final degree of clarity achieved depends on the specific lipid composition used. Some batch-to-batch variation in this step, even among nominally identical lipids, is also possible. Residual haze is due to light scattering by large particles in the suspension, which can be removed by centrifugation. - Incubate the resuspended lipids at RT for 20-60 min.
2. Proteoliposome Formation
NOTE: Several strategies can be employed to insert the detergent-solubilized protein into liposomes. For CLC-ec1 dialysis works well and is therefore the method of choice 6,9,10.
- Add the purified protein to the desired protein-to-lipid (P/L) ratio (expressed in µg of Protein/mg Lipid). The choice of P/L ratio depends on the purpose of the experiment.
- If the goal is to assess the activity of the protein of interest, reconstitute at high P/L’s to maximize the signal, as each liposome contains multiple copies of the protein. To quantify the activity and/or unitary transport rate of the protein, reconstitute in a Poisson dilution regime at low P/L’s, so that each vesicle contains on average 1 protein. See Step 7 and Discussion for a more in depth analysis of when to use each regime.
- To determine the optimal P/L values for the various regimens, perform a complete titration of the activity as a function of protein concentration 8,11-13. However, for any protein for which the molecular weight (MW, expressed in kDa) of the active unit is known the following P/L values can be used as initial guesstimates:
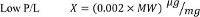
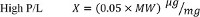
- Load the protein and lipid mixture in a pre-wetted dialysis tube or a cassette. Place the dialysis device in a beaker containing 500-1,000x the volume of the lipid reconstitution buffer (i.e., 1 L buffer for 1 ml of lipid resuspension) under gentle stirring.
- Change buffer 3-4 times at intervals >3 hr. At every solution change, wash the cassette/tubing and the beaker with distilled water and fill the beaker with fresh reconstitution buffer. Include 1-2 O/N intervals for the solution change. The exact number and duration of the solution changes depends on the exact lipid and detergents used.
- After the last step is complete, remove the liposomes from the dialysis vessel, aliquot them in tubes and flash freeze them in liquid N2 or freeze at -80 °C.
3. Recording Set-up
NOTE: The recording set-up (Figure 2A) consists of two chambers (flat bottomed cylinders, ~3-4 ml volume), a Cl– (see below) electrode, a pH meter with an analog or digital electrical output, a digitizer, and a computer with an appropriate acquisition software.
- Connect the Cl– electrode to the pH meter, the output of the pH meter to the digitizer which is connected to the computer. Place the reference electrode in one chamber and the recoding wire in the other (Figure 2B).
NOTE: The polarity of the wires is correct if ΔVCal (see Step 6.4 and 7.2) is positive. - Place the chambers on a stir plate with a stir bar in the recording chamber (Figure 1B). Make sure the stir bar does not touch the electrode.
- Connect the chambers using an agar bridge (Figure 2B) (100 mM KCl and 2% Agarose). Store the Agar bridges in 100 mM KCl.
4. Preparation of the Cl– Electrode
- Take an old and -possibly- non-functioning pH electrode. Break the glass coating to completely reveal the silver wires.
- Carefully remove the coating from the silver wires using a scalpel blade. Clean the wires using copious amounts of H2O and EtOH.
- Place in a saturated solution of FeCl3 (or bleach) until wires are covered with a uniform dark coat of AgCl.
5. Preparation of Unilamellar Vesicles
- The night before the experiments, swell 1 g of Sephadex G-50 beads in 15 ml of external buffer (1 mM KCl, 150 mM K2SO4, 25 mM citric acid, pH 4.5) with gentle shaking (do not stir) for at least 3 hr at RT before use. Each experiment requires ~3 ml of swollen beads. Keep the swollen beads at 4 °C for a few days at most.
- While the liposomes are thawing at RT, pour in each column ~3 ml of swollen G-50 beads. Let them dry by gravity flow at RT; this usually takes 1-2 min.
- Prepare unilamellar vesicles by extruding the proteoliposomes 11 times through a Mini-Extruder using a 0.4 m Teflon cutoff.
- Place the column in a plastic round-bottomed tube. Spin the columns to remove excess solution. Centrifuge for 20-30 sec at 1,400 x g in a clinical centrifuge.
- Discard flow-through, place column in a 13 x 100 mm glass tube and add 100 µl of the extruded vesicles to the column. Spin column for 1 min at 500 x g in a clinical centrifuge.
- Collect ~200 µl of flow-through that will be added to the recording chamber in step 6.5.
6. Efflux Measurement
- Place 2 ml of 100 mM KCl in the reference (ground) chamber and 1.8 ml of the external buffer (1 mM KCl, 150 mM K2SO4, 25 mM citric acid, pH 4.5) to the recording chamber. The slight osmotic imbalance between the internal and external solutions does not affect
- Start the acquisition program. Let the baseline equilibrate, this may take a few minutes.
- Once the signal reaches a stable baseline (the liposomes are ready and the columns dry), start recording (Figure 3A).
- Add 15 µl of a 10 mM KCl solution to calibrate the system (Figure 2A).
- Add liposomes. A small jump might be visible due to incomplete removal of external Cl– from the proteoliposomes (Figure 3A). Wait till the baseline stabilizes.
- Add Valinomycin (1 µl at 1 mg/ml in EtOH) to initiate efflux (Figure 3A).
- Let the efflux run its time course till it plateaus (Figure 3A).
- Add 40 µl of 1.5 M β-octylglucoside (β-OG) prepared in the external buffer to dissolve all liposomes (Figure 3A). End the recording.
7. Data Analysis
- Export the trace file from in an ascii or text format and import it to the analysis program of choice.
- Measure the Voltage at the following points (Figure 3A):
At the beginning of the recording (V0);
After addition of the calibrating pulse (Vcal);
After addition of the liposomes (Vlipo);
At the end of the efflux (Vfin);
After addition of detergent (Vtot);
Baseline the voltages so that V0=0. - Use the Nernst-Plank equation to determine the experimental value of
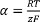 by measuring
by measuring

Where VolCal is the volume of the calibration pulse in µl, 1,800 is the chamber volume at the beginning of the experiment expressed in µl, [Cl]cal/in are the Cl– concentrations of the calibration pulse and of the external buffer in mM and ΔVcal is the jump in mV measured after the calibration pulse.
NOTE: This empirical determination of α serves three purposes: it ensures that the system is responding properly, offers a measure of the consistency of the instrument’s response between experiments as well as to check that no mistakes were made in the solution making. - Calculate the Cl concentrations after each step as follows:
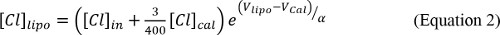


The factor 3/400 comes from the dilution of the 15 µl calibration pulse into the 2,000 µl final volume. - Calculate the changes in [Cl] at each step, ΔCllipo/fin/tot.
- Convert V(t) in Cl(t) with

Directly determine the [Cl] values at the critical points and compare them to the calculated values as an internal control. - Normalize the trace so that the Clrellipo=0 and Clreltot=1

- Fit the efflux time course to the following equation:
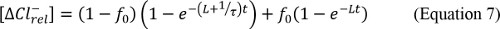
Where L is the rate of Cl– leak that is experimentally determined by measuring Cl– efflux from protein-free liposomes prepared in the same conditions and τ is the time constant of the process. - Calculate v, the initial velocity:

Where ΔClfin is expressed in millimolar and V is the volume of the chamber in μl. - Calculate the transport rate/conductance

Where p is the P/L, MW is the molecular weight of the active complex expressed in kDa
We describe a detailed and robust protocol to measure Cl– transport mediated by purified CLC-ec1, a prokaryotic CLC-type H+/Cl– exchanger, reconstituted in liposomes. A schematic representation of the experiment is shown in Figure 3. Proteoliposomes reconstituted with purified CLC-ec1 and containing high internal Cl– are immersed in a bath solution containing low Cl–. Under these conditions net Cl– efflux is prevented by the buildup of positive charge in the liposome (Figure 3A). Addition of Valinomycin allows K+ to move across the membrane thus shunting the electrical potential and initiating Cl– efflux (Figure 3B). The flux time course is monitored using a Cl–-selective electrode and the total amount of Cl– contained in the vesicles is directly measured by solubilizing them using detergent. From these bulk experiments it is possible to determine properties of the single reconstituted proteins, such as turnover rate, stoichiometry of the active complex and fraction of active protein. Using Equations 7-9 to fit the efflux time course and estimate the unitary transport rate of CLC-ec1 we find that τ(0.2 µg/mg) = 41 sec and f0(0.2 µg/mg) = 0.31, indicating that ~1/3 of the liposomes contains 0 active proteins. From these values we calculate that the unitary transport rate is γ~2,500 Cl– sec-1, in good agreement with published values 8,14.
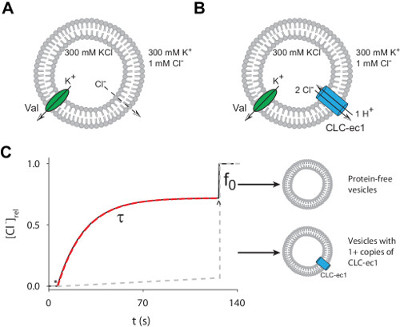
Figure 1: The Cl– efflux assay. (A–B) Schematic representation of the Cl–-efflux assay for protein-free liposomes (A) or CLC-ec1 reconstituted vesicles (B). (C) Simulated time course of ionophore-initiated Cl– efflux from proteoliposomes (black) or protein-free liposomes (gray dashed line). Efflux is initiated by addition of ionophore (*) and terminated by the addition of detergent to dissolve liposomes (^). Red dashed line is an exponential fit to determine the time constant, τ, of Cl– efflux from liposomes containing at least 1 active copy of CLC-ec1 and f0 is the fraction of liposomes containing 0 active proteins. Please click here to view a larger version of this figure.

Figure 2: Recording set up. (A) The recording set up. (B) Close-up of the chambers. Please click here to view a larger version of this figure.
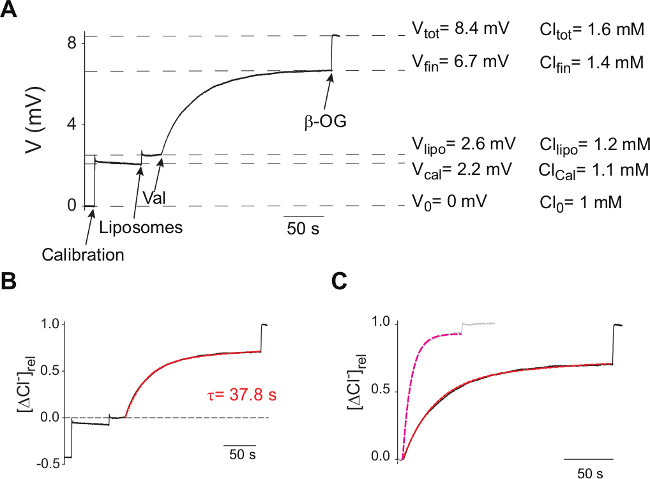
Figure 3: Sample trace and analysis. (A) Typical V(t) traces of efflux experiments from proteoliposomes reconstituted with WT CLC-ec1 at 0.2 μg/mg P/L. Arrows indicate the times of addition of the KCl Calibration pulse, Liposomes, valinomycin and β-OG. Dashed lines indicate the ΔV values at various stages. (B) The V(t) trace is converted into Cl(t) using Equation 5, normalized using Equation 6 and the efflux time course is fit to Equation 7 (red dashed line). (C) Normalized efflux time courses from proteoliposomes reconstituted with WT CLC-ec1 at 0.2 μg/mg P/L (black) or at 5 μg/mg P/L (gray). Dashed lines are best fits of the efflux time courses to Equation 7.
We have described a detailed protocol to measure Cl– transport mediated by purified anion-selective channels or transporters reconstituted in liposomes. The example used was the prokaryotic H+/Cl– exchanger CLC-ec1. However, the methodology can be readily adapted to study channels gated by ligands 12,13,15, voltage 11,12, or sporting different anionic selectivity 15,16 by replacing the Ag:AgCl electrode with one suitable for the ion under consideration. Electrodes selective for ions other than Cl–, such as H+, I– and F–, are commercially available.
A discussion of some of the critical steps and assumptions follow.
Limits of the Poisson dilution assumption
The derivation of the unitary rate (Equation 7-9) is valid only in a Poisson dilution regime, when most liposomes contain only one active protein so that the probability of having multiple copies in a given vesicle is low and the macroscopic efflux time constant of the vesicle population is well approximated by that of liposomes containing a single protein. Indeed, a comparison of the Cl– efflux time course mediated by liposomes reconstituted with CLC-ec1 at low (black, 0.2 µg/mg) and high P/L (gray, 5 µg/mg) (Figure 3C) shows that in the latter case the efflux kinetics become faster and f0 decreases, indicating that a smaller fraction of the total liposomes contains 0 proteins. The total amount of Cl– contained in both vesicle samples was comparable, ~0.7 and ~0.55 µmoles respectively, indicating that the total internal volumes of the vesicles in the two samples was similar and that the amount of protein reconstituted does not affect the size of the vesicles. Fitting the high P/L efflux time course to Equation 7 produces values of τ(5 µg/mg) = 8.8 sec and f0(5 µg/mg) = 0.06, indicating that only ~6% of the vesicles contain no active CLC-ec1 dimers in contrast to the ~31% found when P/L = 0.2 µg/mg. Thus, the estimate of the unitary turnover rate at high P/L is γ~450 Cl– sec-1, nearly 6-fold lower than the one obtained at low P/L and of published data 8,14. Therefore, analysis of efflux time courses from liposomes reconstituted at high P/L’s will underestimate the unitary transport rate of the reconstituted protein. Thus, to accurately determine the unitary transport rate of a novel protein it is of key importance to precisely determine the amount of protein reconstituted and to work at low P/L’s, in a Poisson dilution regime. This is ideally achieved by determining a full protein titration analysis to identify the optimal regime to work at.
Determination of the mass of the active complex
The analysis described above assumes that the stoichiometry of the active complex is known and the reconstituted protein is fully active. However, for new preparations these quantities might not be known a priori. In this case it is possible to directly determine these parameters by measuring the f0’s at different protein to lipid ratios

where p is the protein density,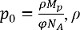 , ρ is the number of liposomes per mass of lipid, NA is Avogadro’s number, MP is the mass of the functional channel complex and φ is the fraction of active proteins. By fitting the experimentally determined f0’s at various protein to lipid ratios it is possible to determine p0, and therefore MP and φ.
, ρ is the number of liposomes per mass of lipid, NA is Avogadro’s number, MP is the mass of the functional channel complex and φ is the fraction of active proteins. By fitting the experimentally determined f0’s at various protein to lipid ratios it is possible to determine p0, and therefore MP and φ.
A finite fraction of liposomes is refractory to incorporation
The derivation of the unitary transport rate assumes that at high P/L’s all liposomes will contain at least one active transport protein. However, in some cases this has been found not to be the case: at high P/L’s a significant fraction of liposomes remains refractory to reconstitution of some proteins 11-13,17. While the origin of this phenomenon are not clear it is conceivable that larger proteins might be excluded from vesicles with smaller radiuses thus reducing the number of available liposomes.
In this case Equation 10 needs to be modified to:
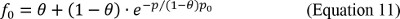
where θ is the fraction of liposomes refractory to protein incorporation. Importantly, ρ and φ are also affected by the reconstitution method since different procedures will have different recoveries for lipids and proteins during liposome reconstitution and formation. Assuming 100% yield for both will lead to a mis-estimation of γ. Therefore, to obtain a precise quantitation of the unitary turnover/conductance it is important to experimentally determine the fraction of lipid and protein lost during reconstitution 8,11.
Orientation of the reconstituted proteins
One of the assumptions made during the analysis is that all reconstituted proteins are functionally equivalent. However, many channels and transporters have preferred directions of transports, a phenomenon known as rectification. This functional asymmetry might lead to an estimation error in the turnover rate. Furthermore, the orientation of the reconstituted proteins is a priori random, so that the measured unitary turnover rate is the weighted average of the protein’s rates when operating forwards or backwards. Several methods can be used to determine the orientation of the reconstituted proteins, for example by measuring the fraction of a tag that is cleavable from the extraliposomal solution or by measuring the sidedness of the reactivity of a single cysteine residue introduced in a soluble-accessible region. Finally, it is possible to circumvent this problem by functionally silencing one of the two populations using sided inhibitors or by selectively activating only proteins in one orientation via the addition of activating compound to side only or by setting the transmembrane voltage to opportune values using K+ gradients, for channels or transporters that are ligand- or voltage-dependent.
Considerations on the formation of tight liposomes
One of the key factors enabling the use of the efflux assay described here is the formation of tight liposomes. Three important variables to be considered to this end are the choice of detergent used to solubilize the protein, the detergent-removal strategy and the choice of lipid composition of the liposomes. In the protocol presented here, CLC-ec1 is solubilized in Decyl-Maltopyranoside (DM), a detergent with a high critical micelle concentration (CMC) value which is therefore easily removed. However, a variety of synthetic and naturally occurring detergents have been used to purify proteins that were subsequently reconstituted in tight liposomes with no effects on the tightness of the vesicles. These detergents have a wide range of CMCs, as high as millimolar (such as DM or Digitonin) and as low as micromolar (such as Dodecyl- Maltopyranoside (DDM) or dioctylpropane-bis-maltopyranoside (DMNG)). It is important to note however, that the lipids used to form the liposomes are solubilized using CHAPS, or another mild detergent, and therefore the hybrid micelles resulting from the mixture of protein and lipids might have chemico-physical properties different from those of the parent detergents. It is therefore possible that in some cases the residual detergent removal that could destabilize the liposomes leading to more pronounced leaks. To circumvent this problem it is advisable to investigate alternative detergent removal strategies, such as biobeads 12,13, spin columns 15 or gel filtration 18. Finally, the lipid composition of the vesicles is an important parameter as specific mixtures might be leaky to different ions in certain conditions. For example, liposomes formed from E. coli polar lipid extract are tight to Cl– at acidic pH’s but become progressively more leaky as the pH increases 19 or liposomes formed from a 3:1 mixture of 1-palmitoyl-2-oleoyl phosphatidyl-ethanolamine (POPE) and 1-palmitoyl-2-oleoyl phosphatidylglycerol (POPG) display a much lower permeability to H+ than those formed from E. coli polar lipid extract 20. It is important to note however that the lipid composition of the vesicles might also affect the activity of the reconstituted protein 13,21,22. Thus, it is important to precisely determine the properties of the protein-free vesicles prepared in each condition and then to assess how this affects the properties of the reconstituted protein.
Generalization to non CLC-type channels and transporters
The efflux assay, as described here, can be directly utilized to determine the single molecule properties of any Cl–-selective channel or transporter, and for example has been used to characterize the properties of the Ca2+-activated Cl– channel TMEM16A 12. We now discuss how to adapt the efflux assay to investigate the properties of transporters or channels that are not Cl– selective and/or have unitary transport rates that are significantly different from the ~103 ion sec-1 of CLC-ec1.
The simplest case is that the protein under study is anion-selective. In this case the only necessary adjustment is to replace the Ag:AgCl electrode with a commercially available one of appropriate selectivity, as was done for the F–-selective Flucs and CLCs 16,23,24 or I– permeable channels such as CFTR 15. If, on the other hand, the reconstituted protein is selective for cations, an ionophore other than valinomycin has to be used to initiate efflux. Given the scarcity of validated Cl– ionophores a useful substitute is a H+ ionophore, such as Carbonyl cyanide 4-(trifluoromethoxy)phenylhydrazone (FCCP). In this case it is important to ensure that the intravesicular pH is maintained constant, for example by increasing the buffering capacity of the internal solution. An alternative strategy is to follow cation efflux through a reconstituted channel or transporter by using protons as a counterion and monitoring H+ transport using a pH meter 25 or a ratiometric pH-sensitive probe 26. However, the indirect nature of these measurements renders quantification of the transport properties of the reconstituted protein difficult. Finally, electrodes selective for a number of cations exist and are commercially available. A third scenario is that the protein under study is permeable to both anions and cations, for example a poorly selective channel 13 or a cation/Cl– cotransporter. In this case, the reconstituted liposomes do not maintain a KCl gradient during the solution exchange process (steps 5.4-5.5) so that they lose their salt content. Therefore in this case all kinetic information is lost. However, the fraction of liposomes containing at least one active protein, f0, can still be determined by adding detergent and measuring the residual trapped Cl– content (step 6.8). This allows for the determination of the molecular mass of the protein by carrying out a protein titration and using Equation 10.
Another aspect to consider is the possibility that unitary transport rate of the reconstituted protein is orders of magnitude different from that of CLC-ec1. For example, most transporters have turnover rates of 1-10 sec-1, 2-3 orders of magnitude lower than CLC-ec1, while most channels conduct ions at 106-107 sec-1, 3-4,000 fold faster than CLC-ec1. For slow transporters the rate of Cl– leakage from protein-free liposomes can become limiting, as its relative weight in the efflux process increases as the protein’s transport rate decreases. In our experience the leak rate varies somewhat between preparations and is usually in the order of a few ion sec-1. Therefore, when working with slow transporters it is advisable to prepare protein-free liposomes side by side to each reconstitution to accurately measure the leak corresponding to each batch of vesicles. The efflux assay has been successfully used to determine the unitary rate of slow transporters with turnover rates around 1-10 ion sec-1, such as a cyanobacterial CLC 27. The converse situation occurs when the reconstituted protein has a high conductance, for example an ion channel. In this case the efflux kinetics are too fast to be resolved as the mixing time (~1-2 sec) and the intrinsic response time of the recording electrode become rate-limiting. In this case, the kinetic information is lost but the mass of the active complex can still be determined 24. It is worth noting that other approaches, such as planar lipid bilayer recordings or patch clamping liposomes are more suitable to study purified ion channels as these techniques directly provide single molecule information with high time resolution.
