The outcome of this protocol depends on the quality of the dissection. This dissection steps must be short (less than 10 to 15 min) and precise (i.e., to avoid damages of the epithelium). The Figure 1 illustrates how an ideal preparation looks like at different magnification levels. At a low magnification under bright field the different cell types (such as knobs of OSNs, supporting cells) are distinguishable (Figure 1A). At the highest magnification level, typically 80X to 160X, in bright field, the dendritic knobs of the OSNs should be clearly distinguishable from the supporting cells (Figure 1B). Under fluorescent light, only the dendritic knobs and cilia of GFP labelled cells are visible (Figure 1C). By comparing the 2 images, the labelled cells can be approached with the recording pipette (Figure 1D).
The temperature of the dissection solution and timing of the dissection are critical. The first part of the dissection, the preparation of the septum (section 3.1.1 to 3.2) should take place within 5 to 10 min in ice-cold solution. The final dissection (3.3) should last less than 5 min at room temperature. In case the dissection lasted for too long, or was performed in dissection solution too warm, the preparation invariably looks rapidly damaged: dendritic knobs are floating above the surface of the epithelium, and resemble dead cells.
Once a seal is reached and the cell opens under the effects of nystatin, typical voltage gated currents can be observed (Figure 2A). The shape and characteristics of these currents can be used to monitor the health of the cell: in a dying cell or if the seal’s quality is decreasing, these currents’ amplitude will decrease. Under the current clamp configuration, action potentials can be recorded either spontaneously (Figure 2B) or by injection of a depolarizing current (Figure 2C). The firing properties induced by current injection characterize the excitability of the recorded neuron. The excitability of different OSNs’ population can be compared (using classical frequency and ISI calculations).
A multibarrel pipette loaded with different odorants and/or different concentrations of odorants can be used to stimulate the cells by pressure ejection. This allows to measure the odorant induced responses. The odorants and concentrations must be chosen depending on the OR expressed in the cell of interest, for example Lyral is the ligand for MOR23, acetophenone for M71, eugenol for mOREG. In the experiment illustrated on the Figure 3, the recorded MOR23-IRES-tauEGFP neurons responded to different concentrations of Lyral, a ligand for MOR23, under current clamp mode (Figure 3A) or under the voltage clamp mode (Figure 3B). In voltage clamp mode recordings, different characteristics can be monitored to quantify the response (maximum amplitude, rise-time, total current elicited, etc.) as performed classically in electrophysiology. Using the maximum amplitude of the odorant-induced response, dose-response can be plotted and fitted using the Hill equation. These results provide information about the encoding properties of each OSN: detection threshold, temporal dynamic, dynamic range and saturation level. Examples of dose-response curves are shown on Figure 3C. All these details can be compared between individual OSNs to measure potential heterogeneity within a population; they can also be compared between OSNs with or without application of a treatment to measure potential modulation and plasticity.

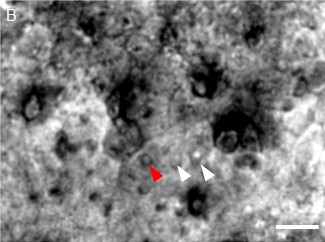
Figure 1: Representative images of a healthy preparation. (A) Intact olfactory epithelium extracted from the nasal cavity of a SR1-IRES-tauGFP transgenic mouse observed under bright field condition at the 40X magnification. OSN dendritic knobs (black arrow heads) are enclosed in a mesh of supporting cells (SC) and Bowman glands (BG). (B) Dendritic knobs of SR1 expressing OSNs observed under bright field condition (white and red arrow heads). (C) The same field as in (B) under fluorescent light showing dendritic knobs of SR1 expressing OSNs. (D) Recording pipette approaching a SR1-expressing OSN under bright field. The red arrowhead represents the same SR1 OSN in (B–D). Scale bar: 5 µm.
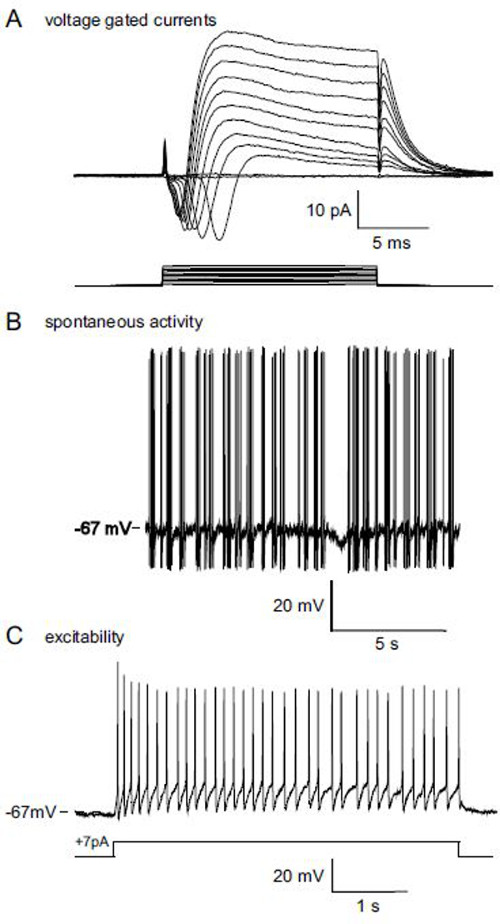
Figure 2: Representative membrane properties results obtained with the protocol: patch-clamp recordings on the dendritic knob of a SR1-IRES-tauGFP OSN. (A) Voltage gated currents elicited by increasing depolarizing steps from the membrane potential -67 mV to +40 mV. (B) Spontaneous activity recorded in the current clamp configuration; action potentials can be observed during the 15 sec recording epoch. (C) Action potentials elicited by a +7 pA excitatory current; this protocol provides information about the excitability of the cell.
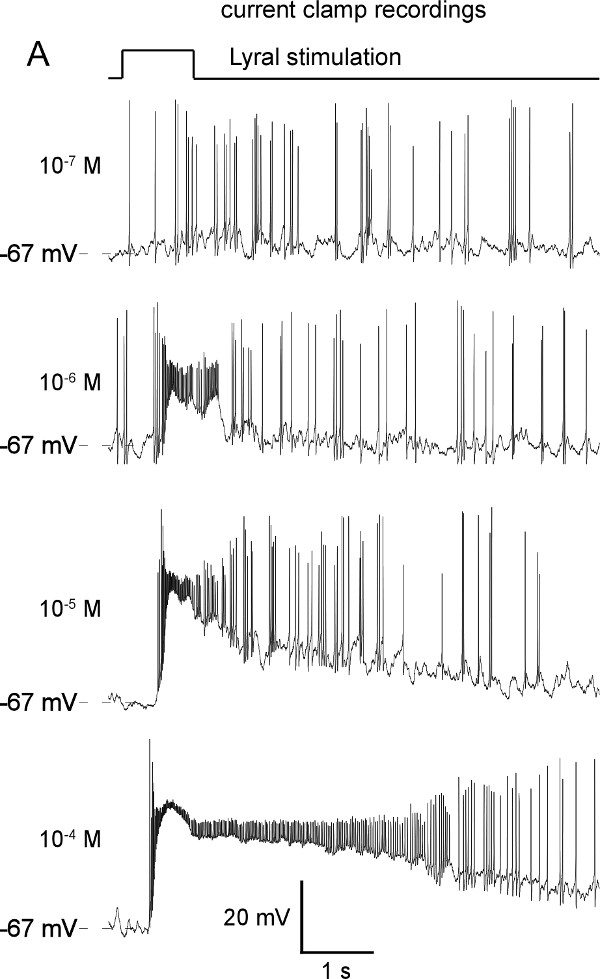
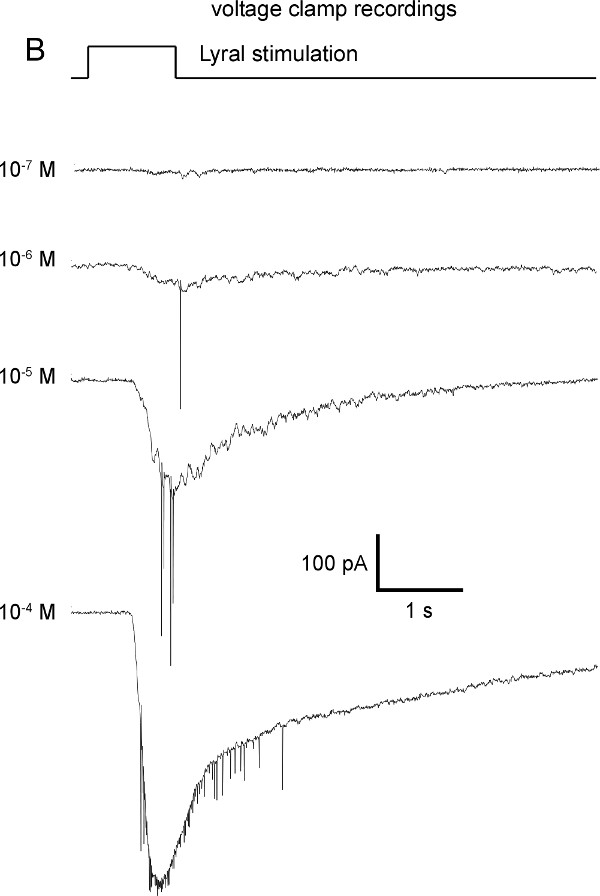
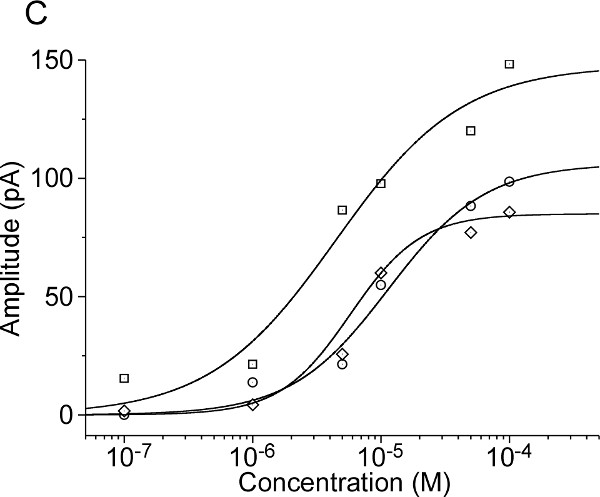
Figure 3: Representative examples of odorant induced responses in a MOR23-IRES-tauEGFP neuron. Increasing concentrations of Lyral, a ligand of the MOR23 receptor, induce increasing responses both in current clamp (A) and in voltage clamp (B). The membrane potential was clamped at -67 mV. (C) Examples of individual dose-response curves acquired on three M71 neurons in response to increasing concentrations of acetophenone and fitted with the Hill equation.
