To understand the regulation of MyoD activity, we undertook the exhaustive characterization of the MyoD complexes using biochemical purification, based on the immunopurification of a double tagged form of MyoD followed by mass spectrometry (MS). The use of HeLa-S3 cell line expressing Flag-HA-tagged MyoD and a control cell line expressing Flag-HA permits to get enough material to purify the MyoD complex by performing double-affinity purification of Flag-HA-MyoD.
We fractionated the cell extracts into cytoplasmic and nuclear fractions, then further fractionated the nuclear fraction to salt-extracted (nuclear salt-extractable, SE) and enriched for nucleosomes (nucleosome-enriched, NE) fractions. TAP-Tag purification from these separated nuclear fractions (Figure 1, Tables 1 and 2) permitted unravelling partners that have relatively low abundance when localized at one specific subnuclear compartment. Furthermore, such a strategy was exploited to uncover partners of unbound (SE) versus DNA-bound (NE) MyoD to get insights on MyoD activity regulation.
For TAP-Tag purification, the Flag-HA tandem epitope system was used. The small hydrophilic Flag and HA epitopes have minimal interference with protein function and are highly accessible for antibody-antigen interaction. Anti-Flag and anti-HA resin-based sequential immunopurification was performed, followed by elution of the immunopurified complexes using Flag and HA peptides. The eluted proteins were then run on an SDS-PAGE to allow all proteins to enter the gel. The pieces of gel containing all purified proteins were cut; the proteins were extracted, trypsin-digested and identified by mass spectrometry (MS).
As shown in Figure 2, MyoD complexes purified from NE fraction have higher enrichment in transcription factors and co-repressors. MS analysis unraveled a series of known partners of MyoD (such as Pbx, Id, E12/E47 (HTF4), BRG1 (SMCA4), MEIS1…) and new partners that were confirmed in the studies generated by this analysis (such as HP1, CBF, MBD3, BAF47 (SNF5/INI1) and all the other SWI/SNF complex subunits…) (Figure 2 and 3)26-28. This sheds light on DNA-bound MyoD partners and possible co-regulators that can establish a repressive-chromatin environment. For example, HP1 proteins, which were identified as MyoD partners by described methodology, are known to bind methylated H3K9 to maintain gene repression and heterochromatin structure. Indeed, HP1 inhibits MyoD transcriptional activity resulting in impaired MyoD target gene expression and muscle terminal differentiation26.
Further fractionation of the SE and NE MyoD complexes on glycerol gradient (as described in29) uncovered the MyoD sub-complexes in the two subnuclear compartments (Figure 1B). In particular, SE MyoD is distributed in three sub-complexes, while the chromatin-bound MyoD belongs mainly to one complex.
Some of the TAP-tag/MS revealed interactors were confirmed by western blot on MyoD complexes. These include the transcription factor CBF, EBB, MTG8R and the SWI/SNF subunit BAF47 (SNF5) (Figure 3A, left) and HP1 proteins (CBX1 and CBX3) (Figure 3A, right). Importantly, since HeLa cells are not muscle cells and do not express MyoD, it is necessary to confirm interactions between newly identified interactors and MyoD in myoblasts (Figure 3 B-D). Notably, for such validation, the total nuclear extracts (without separation on SE and NE) are usually sufficient, which permits reduction of the amount of myoblasts used for sample preparation. The in vitro interaction assays as in26,27, help to further validated these findings. Finally, the functional meanings of these interactions in muscle cells should be further addressed as in26-28.
Taken together, presented data show a global view of ubiquitously expressed MyoD partners and pave the way to further functional studies in a more relevant muscle model.
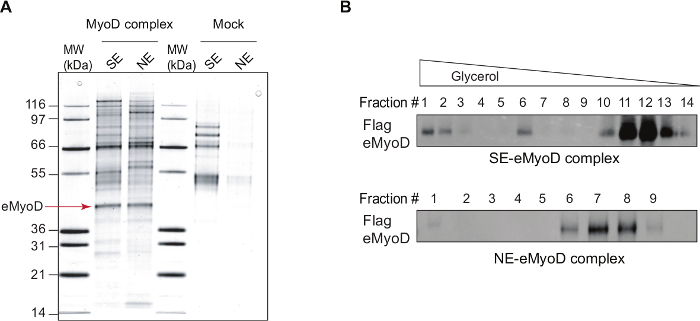
Figure 1: MyoD Complexes Isolated by Tandem Affinity Purification. (A) A silver staining of the double affinity-purified eMyoD complexes isolated from nuclear salt-extractable (SE) or nucleosome-enriched (NE) nuclear fractions of HeLa-S3 cell lines stably expressing Flag-HA-MyoD (MyoD complex) and control cell line (Mock). MW, molecular weight marker in kilo dalton (kDa). Arrow indicates Flag-HA-MyoD (eMyoD). This research was originally published in37. Copyright The American Society for Biochemistry and Molecular Biology. This figure has been modified from26: the lanes with mock purifications and eMyoD complex isolated from SE nuclear fractions are now shown. (B) Double affinity-purified eMyoD complexes as in (A) were fractionated on glycerol gradient ranging from 20% to 41%. Fractions were manually collected, concentrated and analyzed by western blot (WB) using anti-Flag antibodies. Note the presence of several eMyoD-containing sub-complexes in nuclear salt-extractable (SE) fraction. Please click here to view a larger version of this figure.
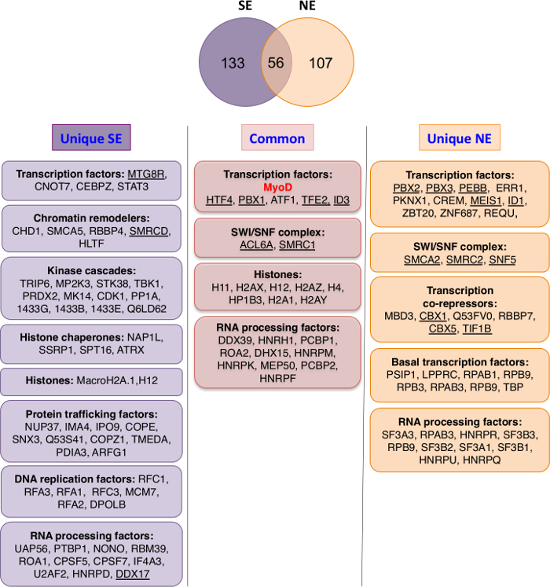
Figure 2: Comparison of Chromatin-bound (nucleosome enriched, NE) versus Nuclear Soluble (nuclear salt extractable, SE) MyoD Partners. Top: Venn diagram showing overlap between eMyoD interactors isolated from nuclear salt-extractable (SE) and nucleosome-enriched (NE) fraction. The ribosomal proteins, translation-initiation factors, DNA repair factors and the tubulin isoforms were excluded from the analysis as they are present in various different data-sets obtained by TAP and considered as non-specific. Bottom: The MyoD interactors found in both SE and NE fractions (common) or specific for one of the fractions (unique) were divided in groups based on their functional annotations. Cytoskeleton-related and other miscellaneous proteins are not depicted. The underlined proteins are MyoD interactors validated either in HeLa and/or in myoblasts by co-immunoprecipitation. Please click here to view a larger version of this figure.
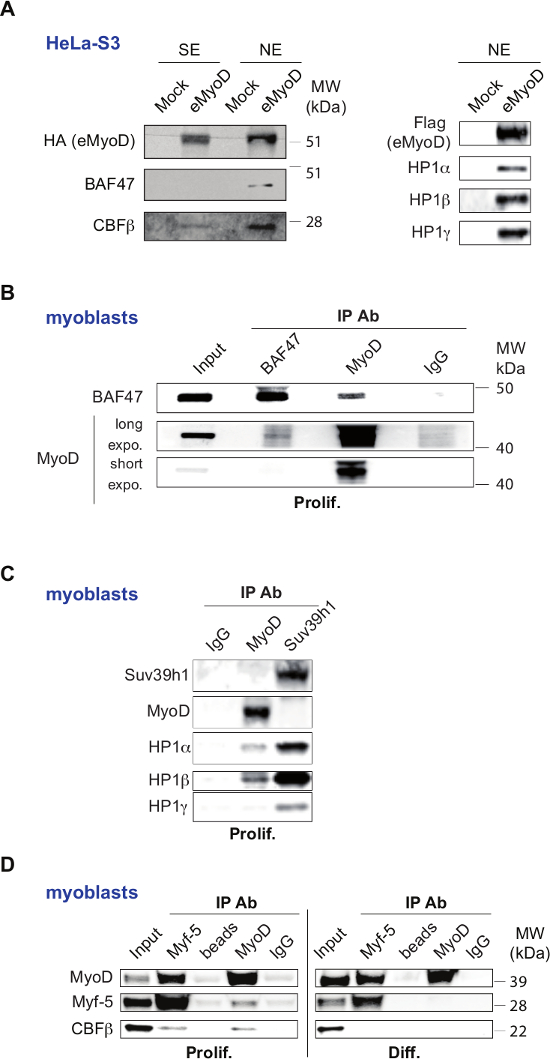
Figure 3: Validation of Selected Set of MyoD Interactors. (A) Validation of MyoD interactors, identified by mass spectrometry, in HeLa-S3 cell line stably expressing Flag-HA-MyoD (eMyoD). Left panel: Western blot analysis of double affinity-purified MyoD complexes isolated from nuclear salt-extractable (SE) or nucleosome-enriched (NE) fractions of HeLa-S3 cell line stably expressing eMyoD or control cell line (Mock) with the indicated antibodies. MW, molecular weight marker. Right panel: Western blot analysis of double affinity-purified MyoD complexes isolated from nuclear nucleosome-enriched fractions of HeLa-S3 cell line stably expressing eMyoD or control cell line (Mock) with indicated antibodies. This panel was originally published in The Journal of Biological Chemistry. Yahi H, Fritsch L, Philipot O, Guasconi V, Souidi M, Robin P, Polesskaya A, Losson R, Harel-Bellan A, Ait-Si-Ali S. J Biol Chem. 2008 Aug 29;283(35):23692-700. doi: 10.1074/jbc.M802647200. Epub 2008 Jul 2. Copyright The American Society for Biochemistry and Molecular Biology. This figure has been modified from26: police type and size was changed and the text was rotated to unify the labeling within the figure. MyoD was labeled as eMyoD to avoid the confusion with endogenous IP presented in the other panels. (B-D) Validation of MyoD interactors in C2C12 mouse myoblasts. (B) Nuclear total extracts from proliferating C2C12 myoblasts were used for immunoprecipitation (IP) with antibodies raised against BAF47 (SNF5) or MyoD, or with control IgG. The resulting precipitates were analyzed by WB with the indicated antibodies. For anti-MyoD antibodies longer (long expo.) and shorter (short expo.) exposure times are shown. Input extracts were loaded to show endogenous protein levels. This panel has been published in28 under the Creative Commons Attribution (CC BY) licence (http://creativecommons.org/licenses/by/4.0/). This figure has been modified from28: police type and size was changed and the test was rotated to unify the labeling within the figure. (C) Nuclear total extracts from proliferating C2C12 myoblasts were used for immunoprecipitation with antibodies raised against MyoD, Suv39h1 (positive control), or control IgG (negative control). The resulting precipitates were then subjected to WB with the indicated antibodies. This panel was originally published in The Journal of Biological Chemistry. Yahi H, Fritsch L, Philipot O, Guasconi V, Souidi M, Robin P, Polesskaya A, Losson R, Harel-Bellan A, Ait-Si-Ali S. J Biol Chem. 2008 Aug 29;283(35):23692-700. doi: 10.1074/jbc.M802647200. Epub 2008 Jul 2. Copyright The American Society for Biochemistry and Molecular Biology. This figure has been modified from26: police type and size was changed and the text was rotated to unify the labeling within the figure. (D) Nuclear total extracts from proliferating (prolif.) and differentiating C2C12 myoblasts (48 hr, indicated as Diff.) were used for immunoprecipitation (IP) with antibodies raised against MyoD and Myf5, or with normal rabbit IgG and with empty beads as negative control. The resulting precipitates were analyzed by WB with the indicated antibodies. Input extracts were loaded to show endogenous protein levels. This panel has been published in27 under the Creative Commons Attribution (CC BY) licence (http://creativecommons.org/licenses/by/4.0/). This figure has been modified from27: police type and size was changed and the text was rotated to unify the labeling within the figure. The panels with the results obtained from proliferating and differentiating C2C12 are separated in two as opposed to the original figure. Please click here to view a larger version of this figure.
Table 1: List of Proteins Identified by MS Analysis in Double Affinity-purified eMyoD Complexes Isolated from Nuclear Salt-extractable Fraction after the Subtraction of the Background Proteins. (Proteins identified by MS in eluates from control cell line were considered as non-specific background.) The data represent the sum of four independent purifications. Please click here to download this file.
Table 2: List of Proteins Identified by MS Analysis in Double Affinity-purified eMyoD Complexes Isolated from Nucleosome-enriched Fraction after the Subtraction of the Background Proteins. The data represent the sum of three independent purifications. Please click here to download this file.
Table 3. Buffer Compositions. Please click here to download this file.
