Cells deficient in coronin A show a defect in early development (Figure 2). In the absence of coronin A cells are unable to form multicellular aggregates, which is the initial step during the developmental cycle of Dictyostelium discoideum. Therefore, coronin A appears to play a role during the early starvation response and/or cAMP signaling. Indeed, the lack of multicellular aggregate formation in the absence of coronin A is accompanied by reduced cAMP signaling 10. However, the application of exogenous cAMP pulses fully restores the wild-type phenotype (Figure 3). This implicates that coronin A, rather than being directly involved in cAMP signaling, functions upstream of the cAMP cascade (Figure 5). When Dictyostelium cells are exposed to starving conditions they induce the secretion of early starvation factors22,23. One of the best characterized factors involved in the early starvation response is conditioned medium factor (CMF)22. Based on our results, we hypothesized that coronin A either regulates the secretion of these early starvation factors or is involved in the signal transduction induced by these factors. In order to be able to differentiate between these two effects we performed conditioned medium experiments. Wild-type or corA-deficient cells were exposed to supernatants from either starved wild-type or corA-deficient cells10 (Figure 4). These supernatants are potent initiators of multicellular aggregate formation and therefore contain factors that initiate downstream developmental signaling pathways, including cAMP signaling. Our results show that supernatant from corA-deficient cells was equally capable to induce development as was the case for supernatant from wild-type cells (Figure 4). This implicates that corA-deficient cells are able to produce and secrete early starvation factors at similar levels to their wild-type controls. However, corA-deficient cells are neither able to respond to their own supernatant nor to supernatant provided from wild-type cells (Figure 4). Therefore, coronin A appears to be involved in signaling pathways induced by early starvation factors, rather than their expression/secretion10 (Figure 5).
In general, to be able to further investigate the signals secreted by starving Dictyostelium cells that induce the developmental cycle, the aggregation assay under submerged conditions is a potent read-out. Cells seeded at higher densities will spontaneously aggregate due to an abundance of all required signals. However, at lower densities only the addition of conditioned medium will induce cellular aggregation (Figure 4)22,24. Therefore, this procedure can be utilized to fractionate conditioned medium and study distinct components for their activity during the early starvation response.
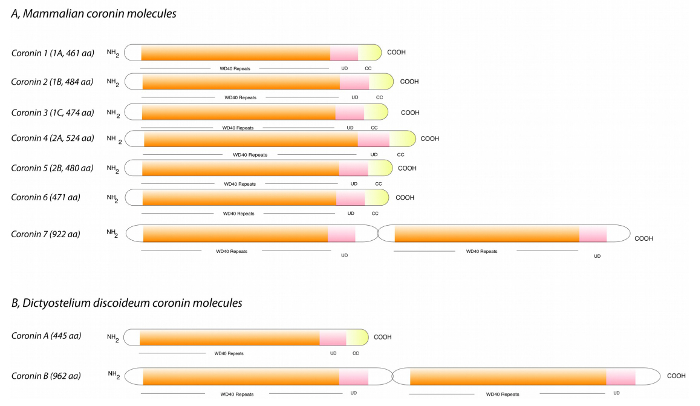
Figure 1: Coronin proteins present in mammals and Dictyostelium discoideum. The coronin family is characterized by the presence of N-terminal WD-repeat regions followed by a unique domain and a C-terminal coiled coil region. While mammals express 7 coronin proteins (A), Dictyostelium discoideum only contains two family members (B), making the risk of redundancy less likely when studying their function. Coronin A is the ortholog of mammalian coronin 1 and coronin B shows a similar domain structure as coronin 7. Abbreviations: CC, coiled-coil domain; UD, unique domain; WD repeats, tryptophan-aspartate repeats. Length is given for mouse sequences (A) in amino acids (aa). Modified from Pieters et al., 201312. Please click here to view a larger version of this figure.
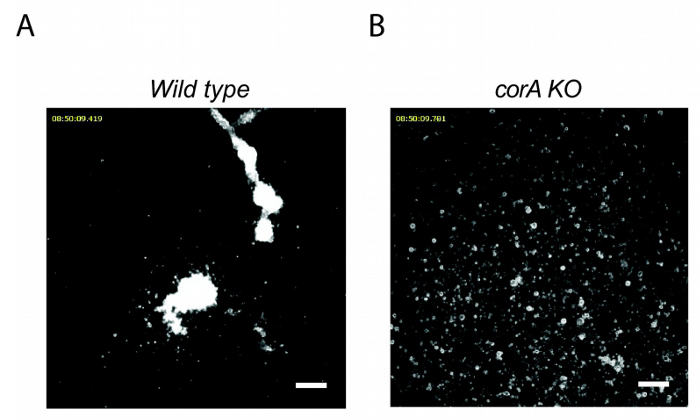
Figure 2: corA–Dictyostelium discoideum are unable to aggregate during the early starvation response. Wild-type (A) or corA-deficient (B) Dictyostelium cells were seeded into multiwell plates at a density of 2 x 105 cells/cm2, starved in BSS, and imaged over a period of 20 hr. Scale bar = 100 µm. Please click here to view a larger version of this figure.
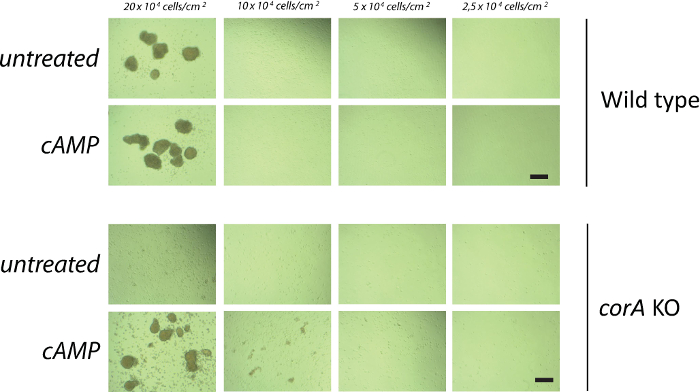
Figure 3: Externally applied cAMP pulses rescue the early developmental defect of corA–Dictyostelium discoideum. Vegetative wild-type (A and B) and corA-deficient cells (C and D) were washed and starved for 2 hr before being pulsed with (B and D) or without (A and C) 50 nM cAMP during 5 hr every 6.5 min in suspension. Cells were then washed, resuspended in BSS, and placed on a Petri dish at a density of 10 x 104 cells/cm2. Images were taken 20 hr after seeding of the cells. Scale bar = 100 µm. Please click here to view a larger version of this figure.
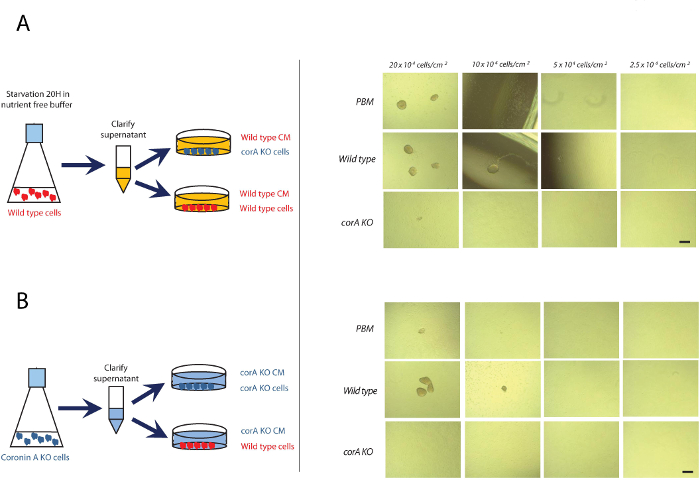
Figure 4: corA-deficient Dictyostelium discoideum are able to produce all required factors for developmental induction but are unable to respond to them. Exponentially growing wild-type and corA-deficient cells were washed in PBM and seeded into multiwell plates at a density of (5, 10, 20, or 40) x 104 cells/cm2, incubated in conditioned medium obtained from wild-type (A) or corA-deficient starving cells (B). The images were taken 16 hr after seeding the cells. Scale bar = 100 µm. Please click here to view a larger version of this figure.
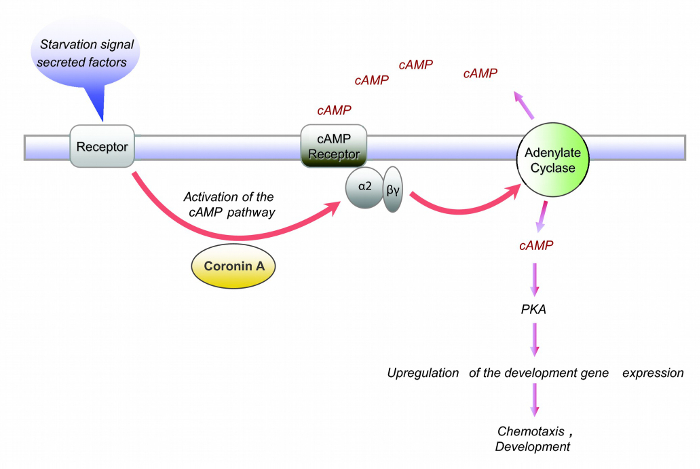
Figure 5: Current model of Coronin A function in Dictyostelium discoideum. Coronin A is involved in the signal transduction of factor(s) secreted during the early starvation response leading to the induction of the cAMP cascade and the progression through the developmental cycle. Modified from Vinet et al., 201410. Please click here to view a larger version of this figure.
