C2C12 cells were transfected with GFP-CUG5 or GFP-CUG200. After drug-resistance selection, stable pools were established, which can be visualized by GFP expression (Figure 1A). Myotube formation in the differentiated myoblasts was detected by myosin heavy chain immunostaining10 (Figure 1B). The quantification of myotube formation demonstrated that fusion indices were decreased from 35.4 ± 4.1% to 2.6 ± 1.1% and myotube areas were decreased from 35.6 ± 2.2% to 2.7 ± 0.8% by abnormal CUG expansion in GFP-CUG200 (Figure 1C). The fusion index and the myotube area were counted via the total number of nuclei in myotubes (≥ 2 nuclei) divided by the total number of nuclei and the percentage of the total image area covered by myotubes, respectively. Real-time RT-PCR10 was conducted to analyze expression for a number of muscle differentiation related genes including MyoD, MyoG, Mef2c and Celf1. Compared to CUG5 cultures, CUG200 increased the expression of Celf1 mRNA in proliferating myoblasts at the beginning of differentiation. Congruent with previous reports, Celf1 upregulation was associated with CUG-expansion in myogenic dystrophy 1 (Figure 1D). Further, Western blotting10 consistently showed Celf1 protein level was elevated (Figure 1E), and CUG200 inhibited the expression of MyoD, MyoG and Mef2c during differentiation (Figure 1D). These results suggest that the CUG-expansion leads to myotube defects and impaired myoblast differentiation.
To study the role of Celf1 in myoblast differentiation, pMSCV- Celf1Flag retroviral vector was constructed to transduce C2C12 cells.10 Puromycin-resistant clones were pooled for differentiation studies. Western blot results showed that the Flag-tagged Celf1 was only present in the pMSCV-Celf1Flag transduced cultures and the expression of Celf1 protein was upregulated (Figure 2A). The formation of myotubes in Celf1-overexpressing cells is rare when compared to the control cultures (Figure 2B), which suggests that the overexpression of Celf1 severely impairs myoblast differentiation.
To determine if Celf1 knockdown rescues the differentiation deficiency in CUG-expansion cells, Celf1 shRNA was delivered to GFP-CUG200 cells by lentiviral vectors. Double resistant clones (G418 and puromycin) were selected and pooled for differentiation studies. The level of endogenous Celf1 protein was markedly reduced in the presence of Celf1 shRNA (Figure 3A). After 6 days of differentiation, the effect of Celf1 shRNA on increasing myotube formation was evident (Figure 3B). Fusion indices and myotube areas were 16.1 ± 3.0% and 15.2 ± 1.2%, respectively, compared to 4.6 ± 0.8% and 5.1 ± 1.3% in control cultures (Figure 3C). Meanwhile, real-time RT-PCR results suggest that the expression of MyoD, MyoG and Mef2c was significantly increased in Celf1 shRNA cells (Figure 3D) supporting Celf1 knockdown rescued myocyte differentiation deficiency.
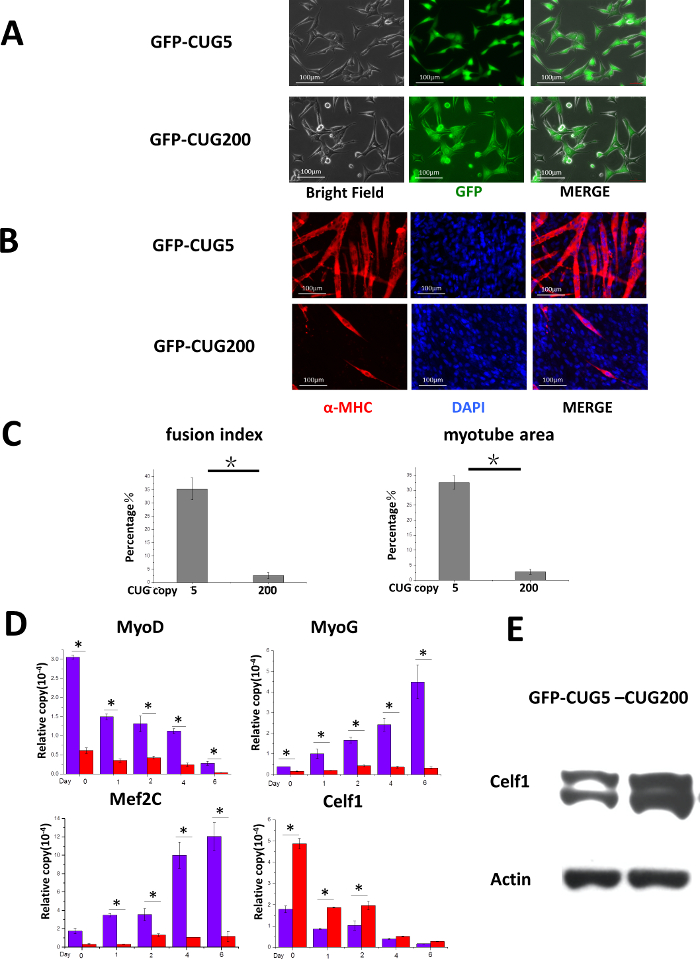
Figure 1. CUG-Expansion Inhibits Myocyte Differentiation in C2C12 Cells. (A) C2C12 cells expressed GFP-CUG5 or GFP-CUG200 ubiquitously after selection. Scale bars are 100 µm. This figure has been modified from our previous paper10. (B) Myotubes were formed in GFP-CUG5 cells but less in GFP-CUG200 cells. Myotubes were visualized by myosin heavy chain (MF-20) immunostaining. Scale bars are 100 µm. (C) GFP-CUG200 cultures significantly reduced fusion index and myotube areas. The error bars represent standard deviations. (D) Real-time RT-PCR analysis of Celf1, transcription factors MyoD, MyoG, and Mef2c expression during myoblast differentiation. mRNA levels were plotted after the GAPDH normalized. The error bars represent standard deviations. Student's T test was performed to compare the tests to controls. n > 3, *P < 0.05. (E) GFP-CUG200 transfection increased Celf1 protein expression. ß-actin is shown as the loading control. Please click here to view a larger version of this figure.
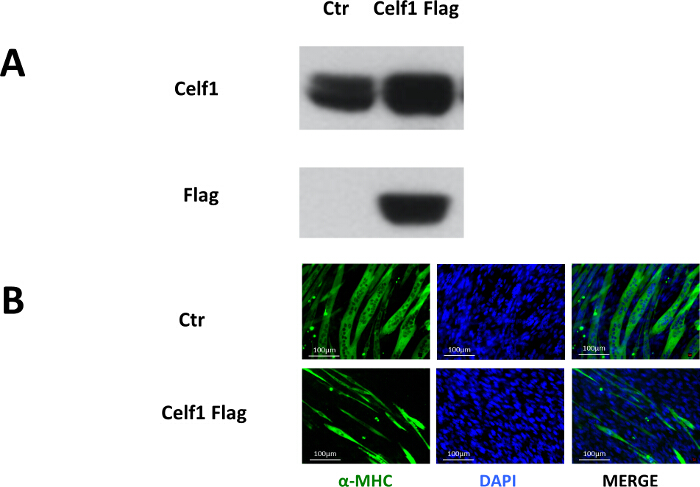
Figure 2. Overexpression of Celf1 Impairs Myoblast Differentiation. (A) Proof of exogenous Celf1Flag expression in C2C12 cells by Western blot. (B) Myotube formation was impaired in Celf1Flag-overexpressing C2C12 myoblasts. Myotubes were visualized by MF-20 immunostaining. Scale bars are 100 µm. Please click here to view a larger version of this figure.
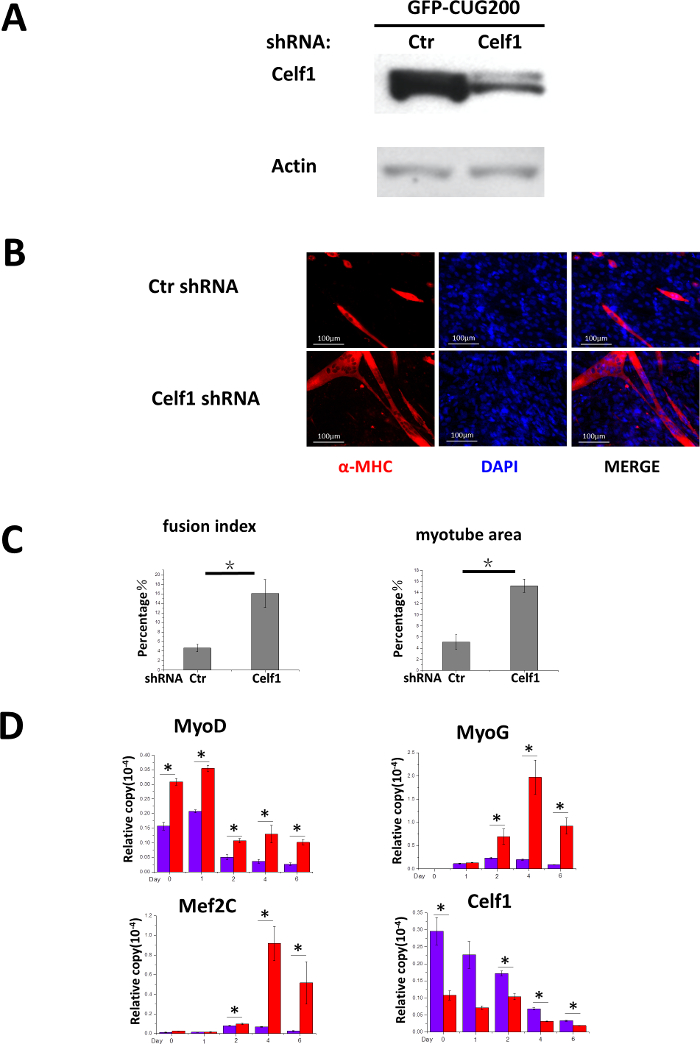
Figure 3. Knockdown of Celf1 Partially Rescues CUG-Expansion-Induced Myoblast Differentiation Deficiency. (A) Proof of Celf1 knockdown by Western blot. (B) Celf1 shRNA induced myotube formation in GFP-CUG200 myoblasts. Myotubes were visualized by MF-20 immunostaining. Scale bars are 100 µm. (C) Celf1 shRNA-rescued cells that had increased fusion index and myotube areas. The error bars represent standard deviations. (D) Real-time RT-PCR analysis of Celf1, transcription factors MyoD, MyoG, and Mef2c expression during differentiation. mRNA level was plotted after normalizing to that of GAPDH. The error bars represent standard deviations. Student's T test was performed to compare the tests to control. n > 3, *P < 0.05. Please click here to view a larger version of this figure.
