To increase the efficiency of siRNA encapsulation within anionic PALETS, the siRNA was mixed with protamine. To determine the best protamine concentration for the siRNA, the siRNA was mixed with different concentrations of protamine, from 1:1 to 2:1 (Figure 3A). There was a 60-65% siRNA encapsulation efficiency in anionic liposomes without the use of protamine. Samples with protamine:siRNA molar ratios from 1:1 to 1.5:1 (133-266 nM) had 80-90% siRNA encapsulation. Molar ratios above 1.5:1 resulted in precipitation of siRNA/protamine complexes. These precipitated complexes were not encapsulated into anionic liposomes. After the addition of protamine to the siRNA there was a slight drop in the concentration of the siRNA, due to the dilution; however, the concentration remained stable after the addition of protamine (Figure 3B). After filtering the liposomes with 50 kDa filters, 90-95% of the siRNA was associated with the liposome retentate, while 5-10% of the siRNA was 'free' in the filtrate. No siRNA was detected in protamine or liposome only samples (Figure 3C).
To determine the effect of LSPCs in vivo, wildtype mice were treated with LSPCs for 24 hr. PrPC expression levels of wildtype mice treated with LSPCs were compared to mice treated with 1x PBS and to PrP knockout (PrPKO) mice (Figure 4). Flow cytometric analysis of PrPC in the brains of mice treated with LSPCs showed a decrease in PrPC levels (Figure 4A and 4B). In one experiment, all mice had 80-90% reduction of PrPC levels, close to PrPC levels that were observed in the PrPKO mice (Figure 4A). Mice treated with LSPCs in two more independent experiments showed a 40-80% reduction of PrPC levels (Figure 4B). Out of the total LSPCs treated mice (n = 14), there was only one mouse had no response to the LSPCs.
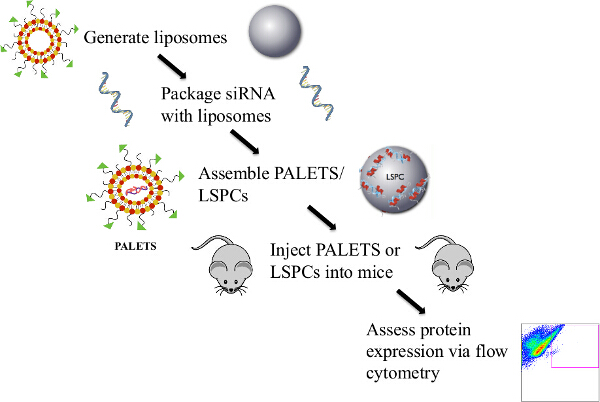
Figure 1: Overview of the Protocol to Generate PALETS/LSPCs and Deliver siRNA Intravenously to the CNS of Mice. Liposomes were generated via the thin lipid film hydration method. The PrPC siRNA was then attached to the liposomes either through an electrostatic interaction or by encapsulation. The PALETS or LSPCs were completed by the addition of the CNS targeting peptide RVG-9r. After assembly, the PALETS/LSPCs were injected into the tail veins of mice, and 24 hr after treatment PrPC expression was assessed via flow cytometry. Please click here to view a larger version of this figure.
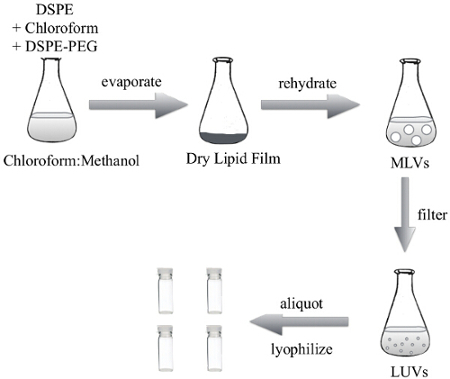
Figure 2: Protocol of the Thin lipid Film Hydration Method to Generate DSPE Liposomes for the PALETS Delivery Vehicle. The lipids were dissolved and mixed in a chloroform:methanol solution, which was evaporated to generate a dry lipid film. The dry lipid film was resuspended in 1x PBS to create multilamellar vesicles (MLVs). MLVs were then uniformly sized using an extruder to generate LUVs. The LUVs can then be used in suspension, or lyophilized in single use aliquots. Please click here to view a larger version of this figure.
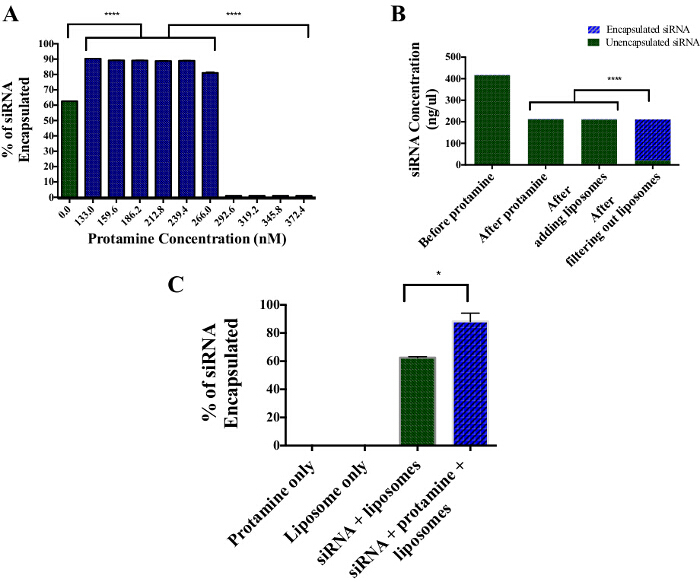
Figure 3: Encapsulation Efficiency of siRNA into DSPE PALETS using Protamine Sulfate. (A) About 60-65% of the siRNA was encapsulated without the addition of protamine. With the addition of protamine, there was a 90% encapsulation efficiency between 1:1 and 1.5:1 protamine:siRNA ratio. (B) After filtering out the liposomes from any free siRNA, 90% of the siRNA was found in the liposomal fraction, whereas 5-10% of unencapsulated siRNA was found in the filtrate. (C) Protamine and liposome only solutions are free of encapsulated siRNA. About 60-65% of siRNA was encapsulated within DSPE liposomes without the use of protamine. With the use of protamine at a concentration of 186.2 nM, about 90% of the siRNA was encapsulated within the DSPE liposomes. Error bars indicate SEM. **** indicates P <0.0001. * indicates P <0.01. Please click here to view a larger version of this figure.
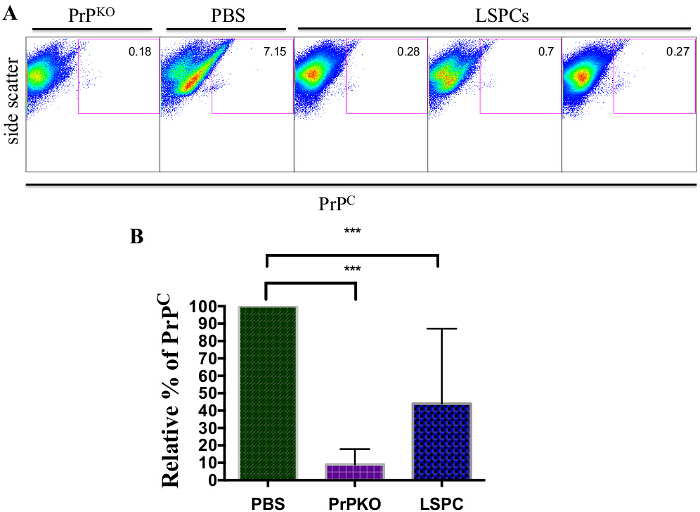
Figure 4: Representative Flow Cytometric Analysis of Brain Cell Suspensions 24 hr after Treatment with LSPCs. LSPCs were injected into the tail veins of wildtype mice and brains were harvested 24 hr later. PBS was used as a treatment control, and a PrPKO mouse was used as a control for PrPC levels. (A) Mice treated with LSPCs showed a significant decrease of PrPC levels compared to the PBS control, which shows wildtype PrPC levels. The LSPCs treated mice showed PrPC levels close to that of a PrPKO mouse. (B) Cumulative data from three independent experiments using the same 24 hr treatment protocol showed the relative amount of PrPC in PBS wildtype and LSPCs treated mice. Across the three experiments, mice treated with LSPCs showed significant decreases in PrPC levels as compared to PBS wildtype controls. Error bars indicate SEM. *** indicates P <0.0003. Please click here to view a larger version of this figure.
