The level of robustness of data obtainable from system described in this report is shown in Figure 2 a-f 38. Through the use of this system the modulation of staphylococcal and fungal interaction with virally infected cells and their effect on each other's adherence can be delineated. These types of studies require microscopic examination of the interaction as shown in Figures 3 and 4 38 in order to determine whether the polymicrobial interaction is occurring on the same cells. In this study differential cell interaction is observed as a result of HSV-modulation of staphylococcal and fungal adherence that is viral species specific.
S. aureus and C. albicans (GT and YF) adhered to the same HSV-uninfected HeLa control cells. This co-localization on cells indicates a lack of physical inference with each other's HeLa cell adherence and that the measured levels of differential adherence measured were likely HSV-mediated (Figure 3A, A1) 38. However, upon HSV-1 or HSV-2 infected HeLa cells no co-localization of staphylococci and C. albicans was observed. (Figures 3B, B1, B2, B3, C1, C2). Using fluorescent microscopy (FITC-conjugated anti-HSV-gD monoclonal antibody) further confirmed that S. aureus did not appear to co-localize with C. albicans nor HSV-1 or HSV-2 (Figures 4A, A1, A2, A3, A4, B1, B2, B3). This co-operation between HSV and Candida extended to both yeast and germ tube forms (Figures 2A-F) of C. albicans. This specificity of association between the triad of microbes reflects the specificity of colonization seen on a much broader scale in the host oronasopharynx mucosa.
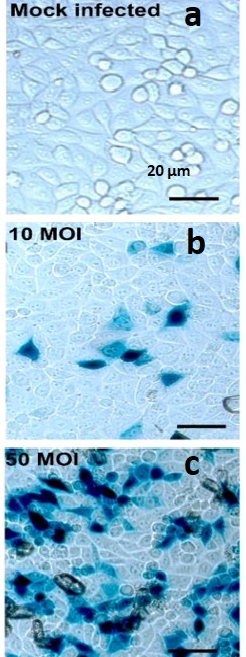
Figure 1. X-gal Staining Pictures of Dosage Dependent HSV-1 Infection in HeLa Cells. HeLa cells infected with HSV-1 at various MOI and X-Gal stained. (A) HeLa cells in well of 96 well plate with X-Gal stained mock-infected HeLa cell control; 20x initial magnification; (B) HeLa cells in well of 96 well plate with X-Gal stained HeLa cell infected with HSV-1 at an multiplicity of infection (MOI) of 10; 20x initial magnification; (C) HeLa cells in well of 96 well plate with X-Gal stained HeLa cell infected with HSV-1 at an multiplicity of infection (MOI) of 50; 20x initial magnification; scale bar applies to all. Please click here to view a larger version of this figure.
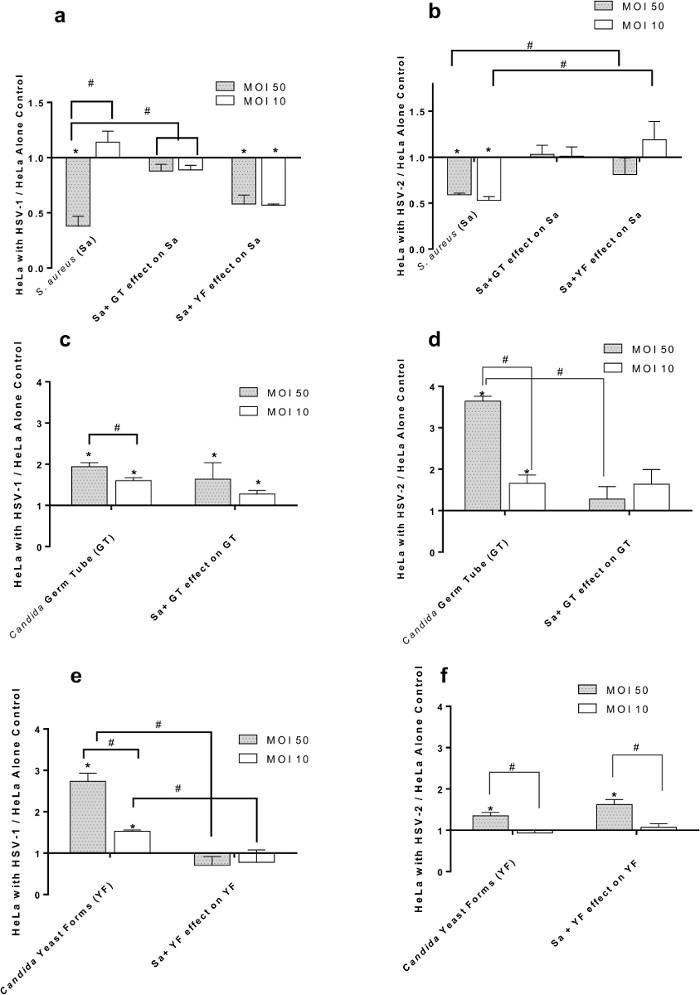
Figure 2. Effect of HSV-1 (panels A, C, E) and HSV-2 (panels B, D, F) at Multiplicities of Infection (MOI) of 50 and 10 on Adherence of S. aureus and/or C. albicans to HeLa Cells. (A) S. aureus (Sa) binding to HSV-1 infected cells in the presence of C. albicans germ tubes (GT) or yeast forms (YF); (B) S. aureus (Sa) binding to HSV-2 infected cells in the presence of C. albicans germ tubes (GT) or yeast forms (YF); (C) C. albicans germ tubes (GT) binding to HSV-1 infected cells in the presence of S. aureus (Sa); (D) C. albicans germ tubes (GT) binding to HSV-2 infected cells in the presence of S. aureus (Sa); (E) C. albicans yeast forms (YF) binding to HSV-1 infected cells in the presence of S. aureus (Sa); (F) C. albicans yeast forms (YF) binding to HSV-2 infected cells in the presence of S. aureus (Sa). All data points are Mean +/- SEM, n= 16 normalized to virus-free control. * = significantly different (p< 0.05) from uninfected HeLa cell control. # = significantly different (p< 0.05) from paired point indicated by bracket; scale bar applies to all.38 Please click here to view a larger version of this figure.
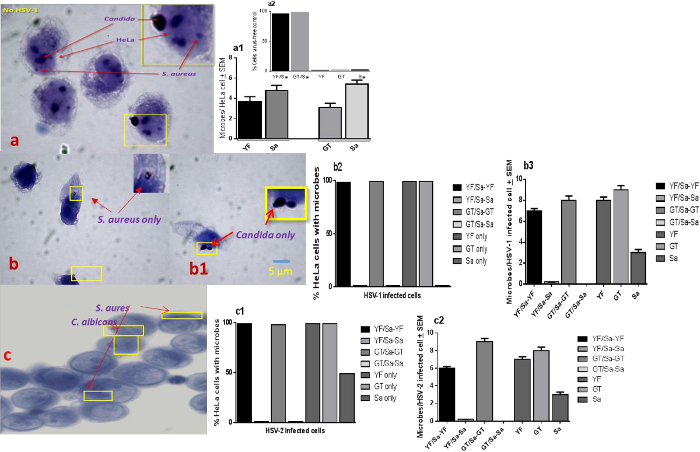
Figure 3. Lack of S. aureus and C. albicans Interactions on HSV-1 and HSV-2 Infected HeLa Cells. HSV-1 and HSV-2 infected (MOI 50) HeLa cell monolayers with S. aureus and C. albicans (5:1 target to cell). For bright field microscopy, cell monolayers were stained with Gram's crystal violet, then examined by light microscopy. Cells that were positive for Candida or S. aureus (100 individual cells per microbe signal/per coverslip) were secondarily scanned for the presence of additional microbe co-localization signals (1,000x initial magnification). (A & A1) S. aureus (Sa) and C. albicans yeast forms (YF) or germ tube forms (GT) co-localize on uninfected HeLa cells; (A2, insert). Percent of HeLa cells with co-localized or individual microbes; (B – B3). Lack of S. aureus and C. albicans co-localization in the presence of HSV-1; (C – C2) Lack of S. aureus and C. albicans co-localization in the presence of HSV-2. Mean ± SEM; scale bar applies to all.38 Please click here to view a larger version of this figure.
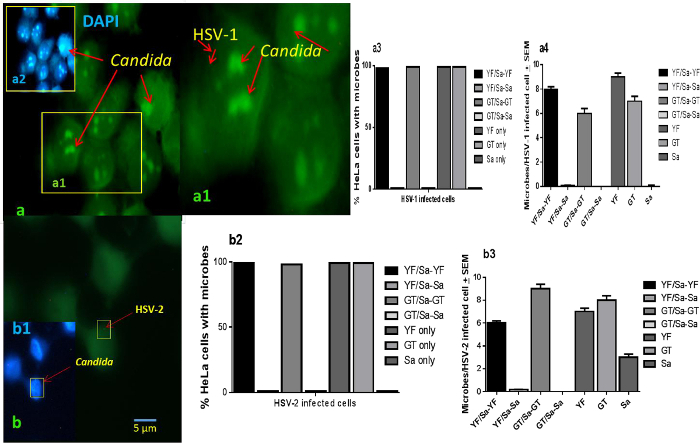
Figure 4. Lack of Co-localization of S. aureus with C. albicans on HSV-1 or HSV-2 Infected HeLa Cells. HSV-infected HeLa cell monolayers challenge with S. aureus and C. albicans (5:1 target to cell) stained (FITC-conjugated anti HSV gD antibody, and DAPI). Pictures are representative of findings from screening of cells that were signal positive for HSV then scanned for Candida and S. aureus (100 individual cells per microbe signal per coverslip) that were then secondarily scanned for the presence of additional microbe co-localization signals (1,000x initial magnification; Nikon). (A – A4) C. albicans (A2, insert; DAPI staining) co-localize with HSV-1; (B – B3) C. albicans co-localized with HSV-2 (B1, insert, C. albicans DAPI staining). Mean ± SEM; scale bar applies to all.38 Please click here to view a larger version of this figure.
| 1 | 2 | 3 | 4 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | ||||||
| Tubes | GT | YF | MSSA | GT/MSSA | YF/MSSA | GT | YF | MSSA | GT/MSSA | YF/MSSA | ||||||
| MEDIA | F | F | MS | MS | F | MS | F | F | F | MS | MS | F | MS | F | ||
| A | ||||||||||||||||
| B | ||||||||||||||||
| C | ||||||||||||||||
| D | ||||||||||||||||
| E | ||||||||||||||||
| F | ||||||||||||||||
| G | ||||||||||||||||
| H | ||||||||||||||||
Table 1.
General Template Determination of Microbial Adherence in Polymicrobial Interactions. GT= Candida albicans germ tube phenotype; YF= C. albicans yeast form phenotype; MSSA= methicillin sensitive Staphylococcus aureus; MS= mannitol salts medium; F= Fungisel medium. Please click here to download this file.
| Plate 1 | 1 | 2 | 3 | 4 | 5 | 6 |
| Control | HeLa cells only | HeLa + GT | HeLa + YF | HeLa + Sa | HeLa + GT+Sa | HeLa + YF + Sa |
| A | ||||||
| B | ||||||
| C | ||||||
| D | ||||||
| Plate 2 | ||||||
| HSV | HeLa cells + HSV | HSV + HeLa + GT | HSV + HeLa + YF | HSV + HeLa + Sa | HSV + HeLa + GT+Sa | HSV + HeLa + YF + Sa |
| A | ||||||
| B | ||||||
| C | ||||||
| D |
Table 2.
General Template for Visual Analysis of Polymicrobial Interactions with HSV-infected and Uninfected HeLa Cells. GT= Candida albicans germ tube phenotype; YF= C. albicans yeast form phenotype; SA= methicillin sensitive Staphylococcus aureus; HSV=herpes simplex virus. Please click here to download this file.
