Here we have demonstrated how to prepare mature Drosophila eggs for ex vivo activation. Eggs expressing a GECI enable imaging of Ca2+ dynamics at egg activation and the beginning of embryonic development (Figure 1). It should be noted that depending on the GCaMP used, specifically the presence of a myristoyl group, results may have slight qualitative differences13 14. We have also demonstrated a role for functional actin during egg activation by the addition of an inhibitor of actin polymerization, cytochalasin D, to the activation buffer (Figure 2) 14.
A requirement for the cytoskeleton at egg activation is conserved. In C. elegans, following fertilization, cytoskeletal components are reorganized to prepare the one-cell embryo for the first asymmetrical division 21,22. In sea urchin, disruption of cytoskeletal re-organization following activation has been shown to affect the cell cycle by preventing contractile ring formation 23. The role of actin in mediating a Ca2+ wave and its downstream function at egg activation in Drosophila remains to be fully elucidated.
In Drosophila, co-visualization of the Ca2+ wave and actin cytoskeleton can be achieved using this ex vivo activation assay. Time series of multiple channels can be acquired and dynamics of the intracellular Ca2+ can be matched with changes in actin organization in the oocyte (Figure 3).
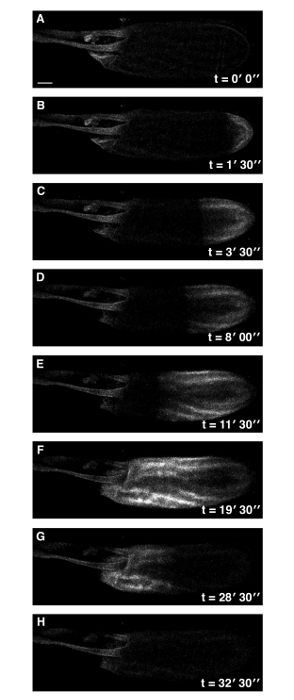
Figure 1: A Single Ca2+ Wave at Drosophila Egg Activation. Time series of ex vivo mature Drosophila egg expressing UASt-myrGCaMP5 following the addition of activation buffer (A-H). The rise in cytoplasmic Ca2+ originates at the posterior pole (B) and propagates (B-F) with an average velocity of approximately 1.5 µm/sec. Initiation of the wave is typically observed within 3 min of the addition of activation buffer. Following activation, the intracellular calcium levels of the mature egg returns to pre-activation levels (G,H). See supplemental movie 1 (total time 33 min). Scale bars = 50 µm. Max projection = 40 µm. Please click here to view a larger version of this figure.
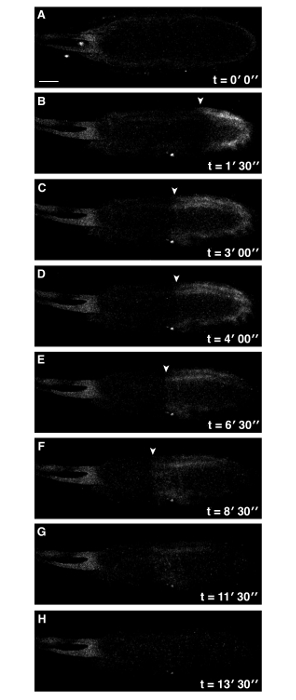
Figure 2: Addition of Cytochalasin D to Activation Buffer Perturbs Ca2+ Wave Propagation. Time series of ex vivo mature Drosophila egg expressing UASt-myrGCaMP5 following addition of activation buffer containing 10 µg/ml cytochalasin D final concentration (A-H). Whilst the Ca2+ wave is initiated at the posterior pole (A), it is compromised and does not reach the anterior of the egg (F) (white arrowheads denote the front of the wave). No further Ca2+ changes were observed over 30 min). See supplemental movie 2 (total time 30 min). Scale bars = 50 µm. Max projection = 40 µm. Please click here to view a larger version of this figure.
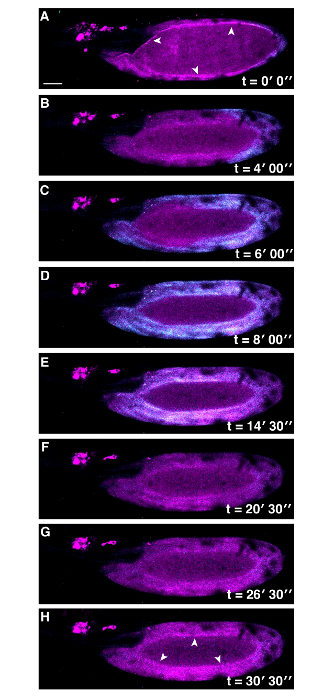
Figure 3: Co-visualization of Actin and Ca2+ at Egg Activation in Drosophila. Time series of ex vivo mature Drosophila egg expressing UASt-myrGCaMP5 (Cyan) and UASp-F-Tractin.tdTomato (Magenta) following addition of activation buffer (A-H). The Ca2+ wave initiates from the posterior pole and recovers as in Figure 1. Actin appears to be changing over time, white arrowheads (A vs H) . The lack of Ca2+ signal detection in the center of the mature egg is due to movement of the sample during image capture. Scale bars = 50 µm. Max projection = 40 µm. Please click here to view a larger version of this figure.
