Field cage study:
Data obtained from the cage experiments showed that the bumble bee colonies had a significant response to fungicide exposure. The fungicide-treated hives produced significantly fewer workers (12.2 ± 3.8, mean ± SE) than the control hives (43.2 ± 11.2, F1,9= 6.8, p = 0.03) (Figure 4). Additionally, the bee biomass of the fungicide-treated hives (0.91 g ± 0.15) was significantly lower than the control hives (2.36 g ± 0.55; F1,9 = 8.3, p = 0.02). This pattern of lowered biomass in fungicide-treated hives was also observed among the mother queens. The queens of the fungicide-treated hives presented with a significantly lower biomass (0.14 g ± 0.04) than the queens of the control hives (0.27 g ± 0.01; Z = 2.5, p = 0.01). However, fungicide exposure did not affect the number of larvae, pupae, and males across the treatments. The biomass of discrete life stages (larvae, pupae, workers, and adult males) and individual weights for the larvae, pupae, and workers did not show any difference across fungicide-treated and control hives.
Lab-based study:
Data obtained from laboratory-based experiments indicated that prior to fungicide exposure, control and fungicide-treated hives had statistically comparable mean worker count (control hives = 28.0 ± 3.1; fungicide-treated hives = 31.67 ± 2.0; n = 3 each) and queen weight (control hives = 0.77 g ± 0.04; fungicide-treated hives = 0.74 g ± 0.01). However, at the end of the study, worker count was significantly higher in control hives (67.67 ± 4.3), compared to fungicide-treated hives (45.33 ± 3.8) (t4 = 3.89, p = 0.01). Worker population increased by ~150% in control hives compared to ~45% in fungicide-treated hives. Similarly, final weight of queen mother remained relatively unchanged in control hives (0.76 g ± 0.02) compared to a ~35% decrease in fungicide-treated hives (0.49 g ± 0.16). Taken together, these results are consistent with previously published results25 and indicate fungicide exposure affected colony fitness as evidenced by fewer worker numbers and reduced queen mother weights (Figure 5).
Metagenomic analysis of pollen-provisions indicated distinct differences between the microbial communities collected from fungicide-treated and control hives (Figure 6). There was a decrease (>95%) in the relative abundance of commonly isolated Streptomycetales, Enterobacteriales in the fungicide-treated hives. Interestingly, both these groups are known for their antifungal activity and role in pollen-preservation30,45 within bumblebee hive environments. Bacterial members of the order Rickettsiales, which includes common pathogens of arthropods46, showed much greater abundance in the fungicide treated hives. Fungicide-treated hives had a lower abundance of fungi belonging to the orders Eurotiales and Sordariales, and a higher abundance of orders Capnodiales and Ascosphaerales compared to control hives (Figure 7). As expected, for both bacteria and fungi, Shannon's diversity index (H) and evenness (E) was lower in fungicide-treated hives compared to controls, although these differences were not statistically significant (bacteria: Hcontrol hives =1.25 ± 0.3, Hfungicide-treated hives = 0.82 ± 0.2, Econtrol hives = 0.46 ± 0.1; Efungicide-treated hives = 0.31 ± 0.1; fungi: Hcontrol hives =0.99 ± 0.3, Hfungicide-treated hives = 0.72 ± 0.4, Econtrol hives = 0.57 ± 0.2; Efungicide-treated hives = 0.42 ± 0.2). Although detailing the metabolic and functional implications of such community shifts was beyond the scope of this study, combined with the colony count, and weight data, these results suggest that fungicide exposure could affect decline in colony health by disrupting the symbiosis between bees and the pollen microbiome. Further investigation into the role of specific microbial groups affecting larval survivorship is recommended to better evaluate the role of the pollen microbiome in sustaining healthy bee populations.

Figure 1: Inside a Bombus impatiens nest. High-resolution image of the workers tending to brood cells. Brood cells can contain several eggs and larvae at any given time. Developing larvae feed on the pollen-provisions periodically introduced into the chamber. Please click here to view a larger version of this figure.

Figure 2: Large field cages erected to house the bee colonies. Each mesh cage (N = 10) was stocked with one commercially purchased Bombus impatiens hive, as well as in-bloom plant species known to attract bees. Flowers were stocked in one corner of the cage and the remaining area was vegetated with oat grass. Please click here to view a larger version of this figure.

Figure 3: Fungicide residue on a recently sprayed sunflower. Field relevant doses of fungicide were sprayed on day 0 and 13 of the experiment. Using a pesticide sprayer, the flowers were evenly coated with a fungicide solution at dusk/evening to avoid direct contact with foraging bees. Please click here to view a larger version of this figure.
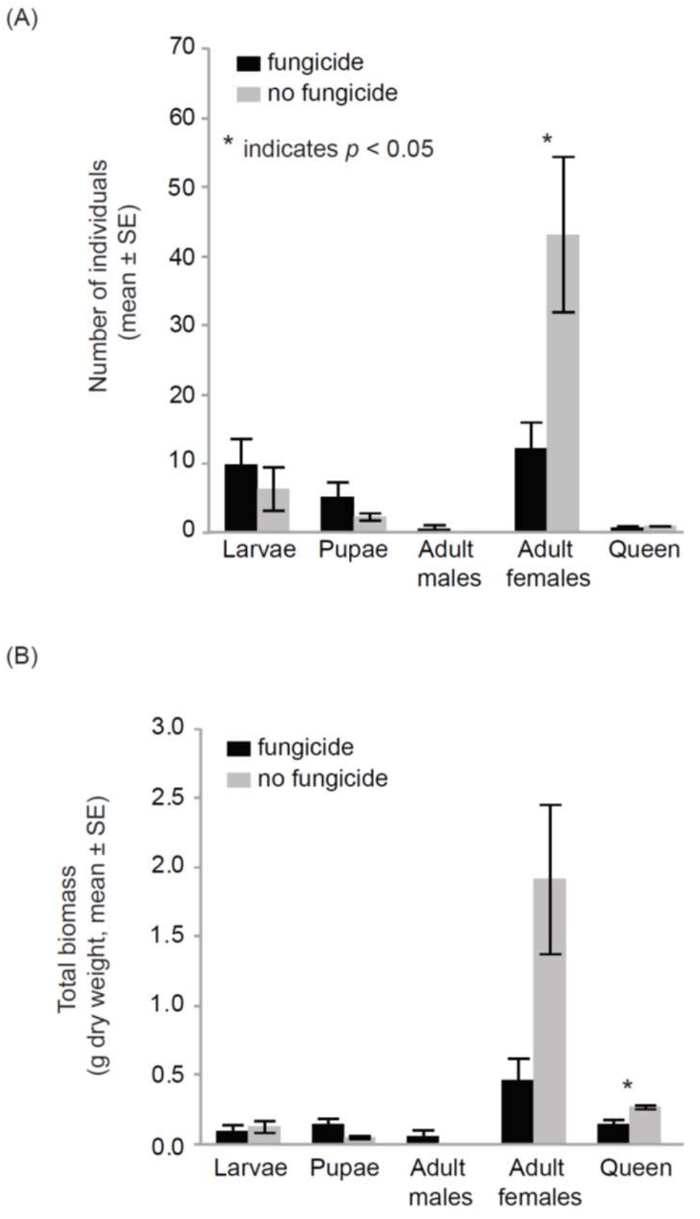
Figure 4: Impacts of fungicide residue on bumble bees in cage experiment. Bumble bee colonies that were exposed to fungicide residues on pollen exhibited significant declines in colony size (reductions in adult female abundance) over the course of a single month. Error bars represent ± 1SE; p <0.05. This Figure was modified from Bernauer et al.25. Please click here to view a larger version of this figure.
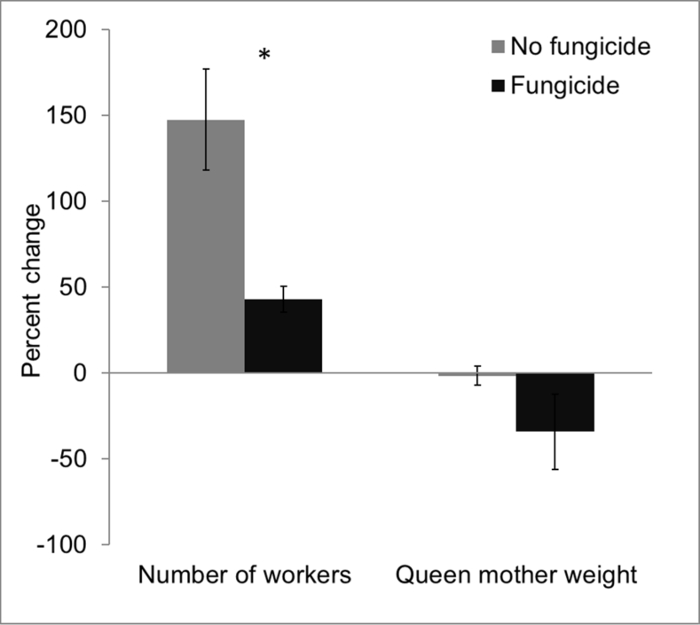
Figure 5: Impacts of fungicide residue on bumble bees in a laboratory-based experiment. Bumble bee colonies that were exposed to fungicide-treated pollen exhibited significant declines in colony size (reductions in adult worker abundance) and decline in queen mother weight over the course of a single month. Error bars represent ± 1SE; p = 0.03. Please click here to view a larger version of this figure.
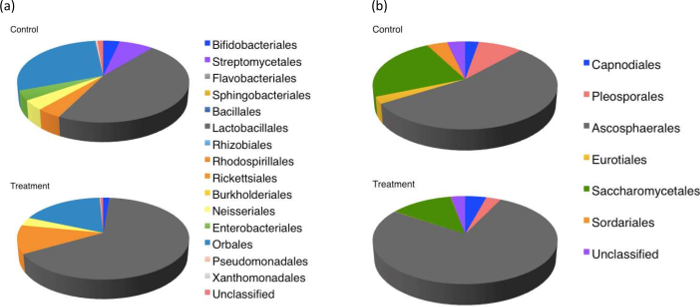
Figure 6: Pie chart of metagenomic classification of fungal and bacterial diversity at the order the rank. Analyses based on (a) 16S and (b) ITS based classification of pollen-provision samples collected from control and fungicide-treated hives. Control hives demonstrated higher diversity and evenness in microbial distribution. Please click here to view a larger version of this figure.
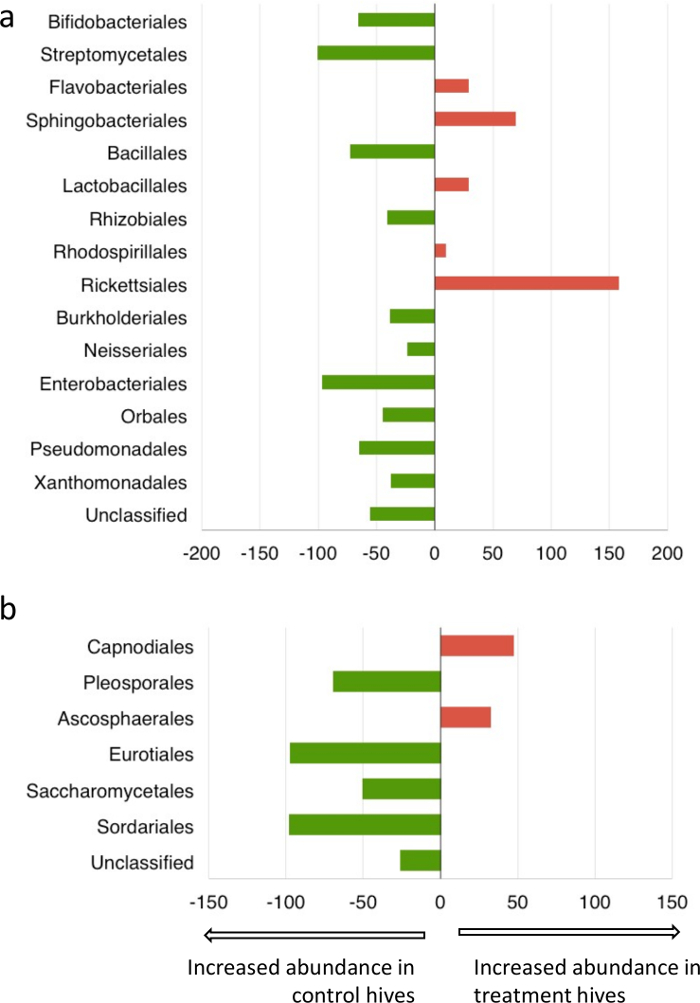
Figure 7: Percent change in order rank relative abundance of bacteria and fungi in response to fungicide exposure. Metagenomic classification of microbial communities using (a) 16S and (b) ITS based primers Please click here to view a larger version of this figure.
| Primer pair | Primer type | Primer sequence | |||
| 28KJ/28B | Plant | GGC GGT AAA TTC CGT CC/ | |||
| CGT CCG TGT TTC AAG ACG | |||||
| ITS1/ ITS5.8 | Fungal | TCC GTA GGT GAA CCT GCG G/ | |||
| GAG ATC CGT TGT TGA AAG TT | |||||
| 16s forward/ reverse | Nested bacterial | 5’- ACACTCTTTCCCTACACGACGCTCTTCCGATCTCCTACGGGNGGCWGCAG-3’/ | |||
| 5’- GTGACTGGAGTTCAGACGTGTGCTCTTCCGATCTGACTACHVGGGTATCTAATCC-3’ | |||||
| ITS1F primer/ ITS4 | Nested fungal | 5’- ACACTCTTTCCCTACACGACGCTCTTCCGATCTTTCGTAGGTGAACCTGCGG – 3’ | |||
| 5’-GTGACTGGAGTTCAGACGTGTGCTCTTCCGATCTTCCTCCGCTTATTGATATGC – 3’ | |||||
| Adapter primers | 5’-AATGATACGGCGACCACCGAGATCTACAC[55555555] ACACTCTTTCCCTACACGACGCTCTTCCGATCT-3’ |
||||
| 5’-CAAGCAGAAGACGGCATACGAGAT[77777777] GTGACTGGAGTTCAGACGTGTGCTCTTCCGATCT -3’ |
|||||
Table 1: List of primer pairs and primer sequences used in DNA amplification. See text for references.
