In the sea urchin embryo we have shown that 3 different Wnt signaling branches (Wnt/β-catenin, Wnt/JNK, and Wnt/PKC)4,25 interact to form a Wnt signaling network that governs anterior-posterior (AP) patterning. One of the most important consequences of these signaling events is that the initial broadly expressed anterior neuroectoderm (ANE) GRN becomes restricted to a small territory around the anterior pole by the beginning of gastrulation (24 hpf in S. purpuratus). These results indicate that Wnt/β-catenin signaling prevents ANE gene activation in the posterior half of the embryo by the 32-cell stage. Then, this pathway relays a signal to the non-canonical Wnt/JNK signaling pathway that progressively down regulates the ANE GRN in the anterior half of the embryo between the 60-cell stage and early gastrulation. Finally, a third non-canonical Wnt pathway, Wnt/PKC, antagonizes the Wnt/JNK signaling pathway and prevents it from eliminating ANE specification around the anterior pole (Figure 1A)4.
We use the spatiotemporal expression of foxq2 to assay for the activity of each of the Wnt signaling branches during AP patterning because it is one of the first two genes activated in the ANE GRN and it is easily assessed by in situ hybridization because of its robust expression26. If any individual Wnt signaling branch is perturbed, then there are clear expression phenotypes that indicate which pathway is involved: 1) In the absence of Wnt/β-catenin signaling a broad maternal regulatory mechanism (Figure 1A) activates foxq2 expression throughout the entire embryo (Figure 1Bb); 2) In the absence of Wnt/JNK signaling foxq2 is expressed throughout the anterior half of the embryo (Figure 1Bc), but it is still down regulated in the posterior half due to the activity of Wnt/β-catenin signaling (Figure 1A); In the absence of Wnt/PKC signaling foxq2 expression is completely down regulated throughout the embryo (Figure 1Bd), because the Wnt/β-catenin and Wnt/JNK signaling pathways are up regulated4,25. Thus, we have developed an assay that we have termed the foxq2 transcriptional readout system which, when combined with our systematic workflow (Figure 2), allows us to efficiently identify and test whether a gene of interest is involved in one or more of the Wnt signaling branches.
Using the methodology and foxq2 readout system presented here, we have identified several putative extracellular or intracellular molecules likely involved in the Wnt network governing AP axis specification, four of which are presented in Figure 3. Embryos injected with morpholinos designed to knockdown the expression of either the transcription factor, ATF2, or the secreted extracellular Wnt modulator, Wif-1, showed an expansion of foxq2 expression toward the posterior pole of the embryo at early gastrulation stages (Figure 3A – C). These results mimic the phenotypes observed when members of the Wnt/JNK signaling pathway are knocked down4,25, suggesting that they are members of this signaling branch that is necessary to down regulate ANE GRN expression in the anterior half of the embryo. In contrast, foxq2 expression was eliminated when we knocked down the expression of the transcription factor, NFAT (Figure 3D), and the secreted extracellular modulator, sFRP3/4 (Figure 3E), suggesting that these molecules are involved in the Wnt/PKC pathway that is necessary to antagonize the down regulation of the ANE GRN by Wnt/JNK signaling. Based on these rapidly obtained results we are now able to perform more detailed functional analyses that will place these factors within the evolving Wnt signaling network involved in the GRN that governs AP patterning in the sea urchin embryo.
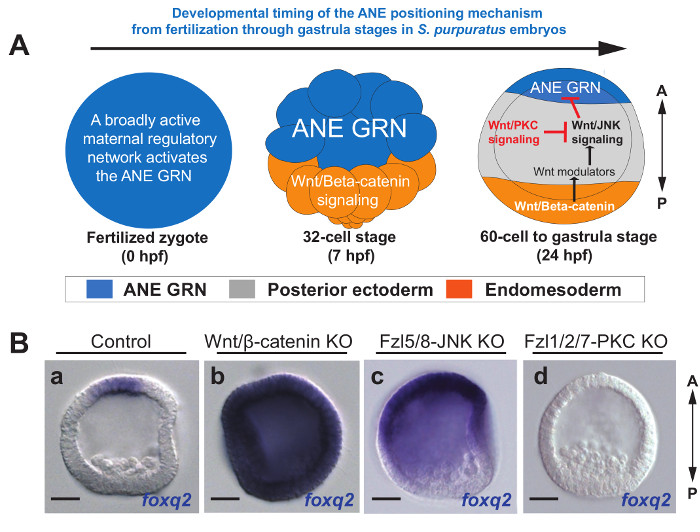
Figure 1. The Model for AP Specification and Patterning in the Sea Urchin and the foxq2 Transcriptional Readout System. (A) The progressive down regulation of the ANE GRN to a territory around the anterior pole by the Wnt signaling network detailed in the text and in4,9. (B) A transcriptional readout system showing foxq2 expression in the indicated Wnt pathway knockdowns (KO). Scale bar = 20 μm. Please click here to view a larger version of this figure.
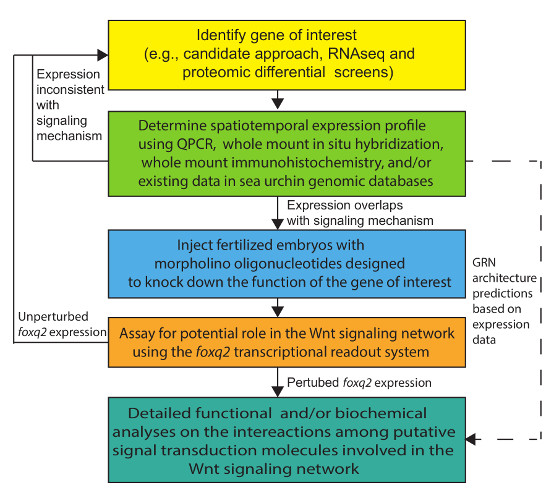
Figure 2. An Experimental Flow Diagram for Efficient Wnt Network Analysis in the Sea Urchin Embryo. Please click here to view a larger version of this figure.
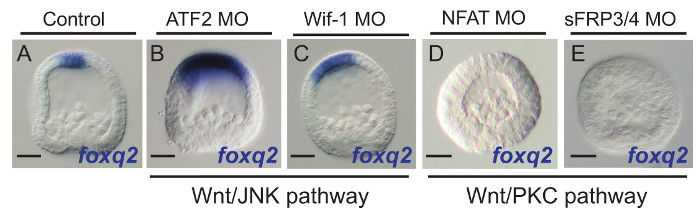
Figure 3. Signal Transduction Molecules Involved in the Wnt Network Governing AP Axis Specification and Patterning Identified Using the foxq2 Transcriptional Readout System. (A, B, C) Morpholino knockdowns suggest that an intracellular signaling modulator, ATF2, and secreted extracellular modulator, Wif-1, are potential players of the Wnt/JNK signaling pathway. (A, D, E) Knockdown experiments indicated that NFAT, an intracellular signaling modulator, and sFRP3/4, a secreted Wnt signaling modulator, are involved in Fzl1/2/7-PKC signaling. Scale bar = 20 μm. Please click here to view a larger version of this figure.<!–
