The key to a successful preparation of PMVs depends heavily on the correct assembly of the filter unit (Figure 1), which can be tested by pushing 1 mL of PBS through the membrane. If leakage occurs, reassemble the filter unit and test again. However, the leakage can only be tested reliably when cells are pushed through the membrane. If only a few PMVs are detected under a regular microscope using the 10X objective, or if the size of the PMVs is mostly around 1 µm, that would indicate that most of the cells are trapped inside the unit due to incorrect assembly. PMVs with a size in the range of about 3 µm in diameter were predominant when examined under a phase contrast microscope using the 20X objective (Figure 2). Since PMVs of nanometer-size were not easily visualized, the percentage of large PMVs in terms of number was low. It was found to be helpful to press the syringe quickly to reduce the amount of membrane debris, which apparently fails to form spherical PMVs. Furthermore, PMVs of smaller sizes (less than 1 µm) were found in the first extrusion medium, with larger PMVs recovered in the second extrusion.
The most unique aspect of the PMVs was the enclosing of cellular components aside from the nucleus12. BMSCs were transfected with an expression cassette of EGFP for 48 h before being used for PMV preparation. The results show that cytoplasmically localized EGFP protein can be encapsulated in PMVs (Figure 3A). The enclosure of mitochondria was evidenced by the staining of mitochondrial dye, showing that a large fraction of, but not all, PMVs contain green fluorescent dots, indicative of mitochondria (Figure 3). Some of the PMVs, even those with a diameter as large as 3 µm, did not show evidence of containing mitochondria, suggesting that some mechanical constraints might prevent the encapsulation of mitochondria into PMVs. The membrane potential of mitochondria was detected by JC-1 staining, which emits red fluorescence when mitochondria have a normal potential. The result show that red fluorescence indeed was detected in PMVs (Figure 3), while the green fluorescence was not detectable (data not shown), suggesting that the mitochondria within the PMVs were functional.
A concentration process for PMVs is needed, especially for in vivo application to reduce the volume as well as to remove the cellular components released during extrusion. After centrifugation at 1,200 x g for 5 min at room temperature, very few PMVs of smaller size were found in the supernatant. To facilitate the resuspension of the pellet, a 2-mL round-bottom tube was used for the centrifugation. However, most of the PMVs were aggregated in the pellet, especially PMVs of smaller sizes (Figure 4). This is probably due to adhesion molecules on the membrane. The medium was therefore supplemented with PEI (2 µg/mL), whose concentration was tailored to have no noticeable cytotoxicity. The positive charge of PEI attached to the cell membrane prevented the aggregation of PMVs during centrifugation (Figure 4).
Finally, the membranes of BMSCs were labeled with CM-DiI dye before the preparation of the PMVs (Figure 5). After concentration, PMVs were injected into the gastrocnemius muscles of mice, which were harvested 12 h post-operation and examined under a confocal microscope. Frozen sections of the gastrocnemius muscle were examined for the presence of red fluorescence, which was detected within the myofiber (Figure 5), demonstrating that PMVs were delivered to the cytoplasm, probably via endocytosis. Of note, red fluorescence was found around the periphery of the myofiber (data not shown), indicating that a large fraction of PMVs were either not endocytosed or were just at the beginning of endocytosis at 12 h post-injection.

Figure 1: Assembly of the filter unit. (A) Commercially available polycarbonate membranes. (B) Track-etched pores of 3 µm in diameter, viewed under an optical microscope with a 20X objective. Scale bar = 50 µm. (C) The membrane was placed on top of the supporting matrix of a disposable filter unit. (D) The assembled filter unit with a membrane inside. Please click here to view a larger version of this figure.
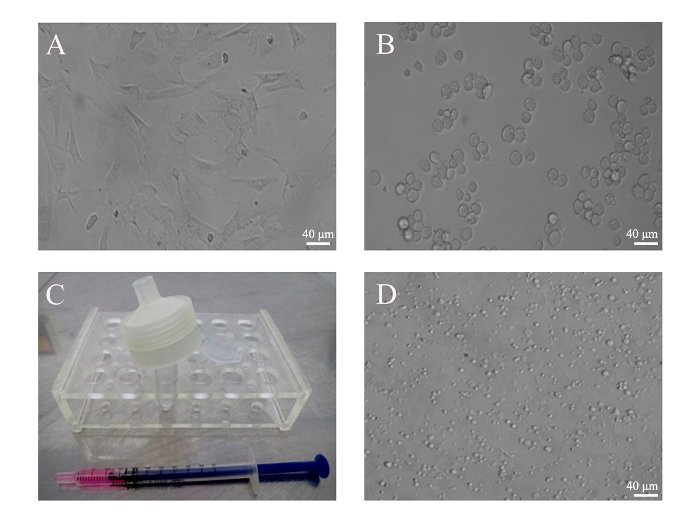
Figure 2: Generation of PMVs from BMSCs. (A) BMSCs from mice were cultivated in a 6-well plate. (B) BMSCs (5 x 105) were harvested by trypsin digestion and monodispersed in 500 µL of culture medium. Images of attached and resuspended BMSCs were taken under an optical microscope with a 20X objective. (C) Cells were loaded into an insulin syringe before being attached to the filter unit. (D) PMVs generated from BMSCs were loaded into a 35 mm glass-bottom dish and examined under an inverted phase contrast microscope using a 20X objective lens. Scale bars = 40 µm. Please click here to view a larger version of this figure.
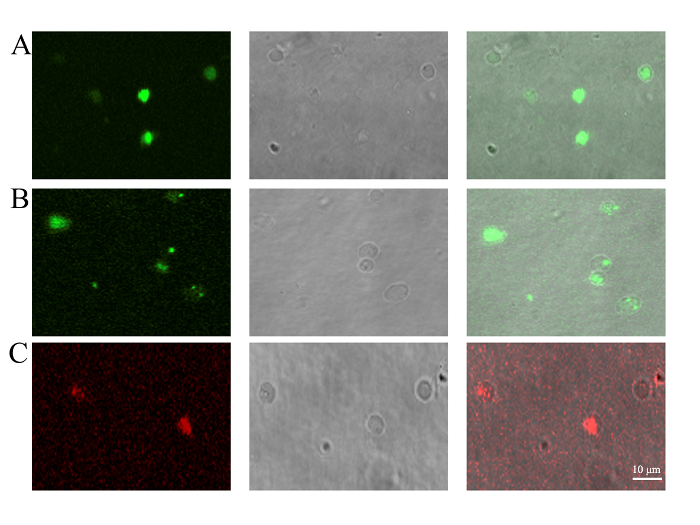
Figure 3: Detection of PMV content by confocal microscopy. PMVs were generated from (A) BMSCs transfected with pEGFP-N1 for 48 h to trace the cytoplasmically localized EGFP protein, (B) BMSCs stained with mitochondrial dye (1 µM) for 30 min to trace the existence of mitochondria, or (C) BMSCs staining with JC-1 (10 µg/mL) for 30 min to confirm the membrane potential of mitochondria. PMVs generated from BMSCs were loaded onto a 35 mm glass-bottom dish and examined by confocal microscopy using a 100X oil objective lens. Scale bars = 10 µm. Please click here to view a larger version of this figure.
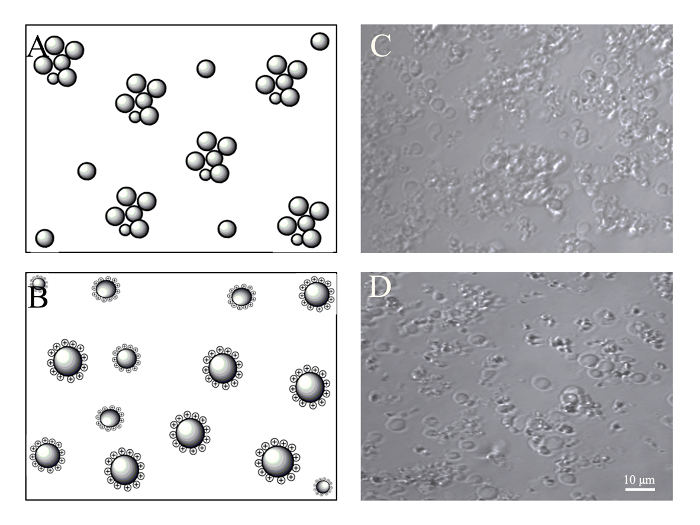
Figure 4: Concentration of PMVs by centrifugation. BMSCs (5 x 105) were harvested by trypsin digestion and resuspended in 0.5 mL of culture medium, which was supplemented with or without 2 µg/mL of PEI. Generated PMVs were spun at 1,200 g for 5 min, resuspended in 100 µL of culture medium, and examined under a confocal microscope with a 40X oil objective. (A) Schematic of PMV aggregation probably caused by the interaction of adhesion molecules after centrifugation. (B) Schematic of PMVs maintaining monodispersion after being charged with PEI. (C) Aggregated PMVs after centrifugation. (D) PMVs show less aggregation after centrifugation due to positive charging with PEI. Scale bars = 10 µm. Please click here to view a larger version of this figure.
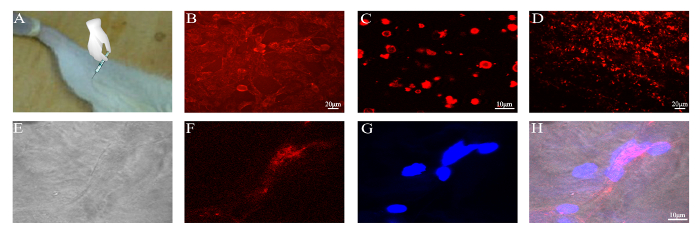
Figure 5: Delivery of CM-DiI-labeled PMVs to the gastrocnemius muscle. PMVs were generated from 5 x 105 BMSCs, concentrated by centrifugation into a volume of 100 µL, and injected slowly into the gastrocnemius muscle of a BALB/c mouse. (A) Schematic of PMV injection. (B) BMSCs labeled with CM-DiI; Scale bar = 20 µm. (C) PMVs generated from CM-DiI-labeled BMSCs and examined by confocal microscopy; Scale bar = 10 µm. (D) Gastrocnemius muscle harvested 12 h post-operation; Scale bar = 20 µm. (E–H) Frozen section of the gastrocnemius muscle examined by confocal microscopy; Scale bar = 10 µm. Please click here to view a larger version of this figure.
