We have developed a three-way strategy that maintains a healthy population for both visual and behavioral studies (summarized in Figure 1). Our method starts with collection of hoverflies from the wild (Step 1, Figure 1). In our lab, hoverflies are housed in insect cages or plastic bags, under an artificial hibernation cycle (Step 2, Figure 1), substantially prolonging their life span. For increased numbers, offspring can be reared from wild-mated females (Step 3, Figure 1).
We have found that catching vast numbers of wild hoverflies is a time intensive endeavor, even when environmental conditions are favorable. In contrast, the successful rearing of mature larvae harvested from the manure pits of cattle farms is a far more efficient way to source large numbers of wild hoverflies (Step 1, Figure 1), with us collecting up to 700 larvae in 0.03 m3 of manure. Additionally, our techniques to rear eggs laid by captured gravid females have proved to be successful (Step 3, Figure 1). Females captured in a Mediterranean climate (Adelaide) during the autumn and winter months laid several batches of eggs, with 24 clusters observed from 19 females in a period of 20 weeks. Of these egg batches, 10 were placed into water, all of which were fertile and resulted in the hatching of larvae. 3 groups of larvae were then taken beyond this point, and placed in the rabbit feces slurry, resulting in 163 ± 34 (mean ± SD, N=3) emerged hoverflies, with no observed gender bias (Figure 2).
The health of these lab reared hoverflies was determined by a comparison of the weight and locomotor activity of female hoverflies compared to field captured individuals. General locomotor activity was assessed using a Locomotor Activity Monitoring system (LAMS), as described previously39. No significant differences in weight (Figure 3A) or activity (Figure 3B) were observed between lab reared and wild caught hoverflies after 4 months in captivity under our artificial hibernation cycle. When E. tenax were maintained in the laboratory without the use of an artificial hibernation cycle we saw a significant decrease in longevity, with a lifespan of 2.5 – 3 months (73 ± 7 days for 5 females and 79 ± 4 days for 11 males). When the hoverflies were kept in artificial hibernation they could live in excess of 12 months.
Additionally, the effect of long term maintenance, using our described methods, was further assessed by a comparison of weights over time for both sexes of lab reared hoverflies. We observed a significant increase in weight over a period of 4 months for both sexes, with females consistently weighing more than their male counterparts (p < 0.0001, two-way ANOVA, N=12, Figure 4).
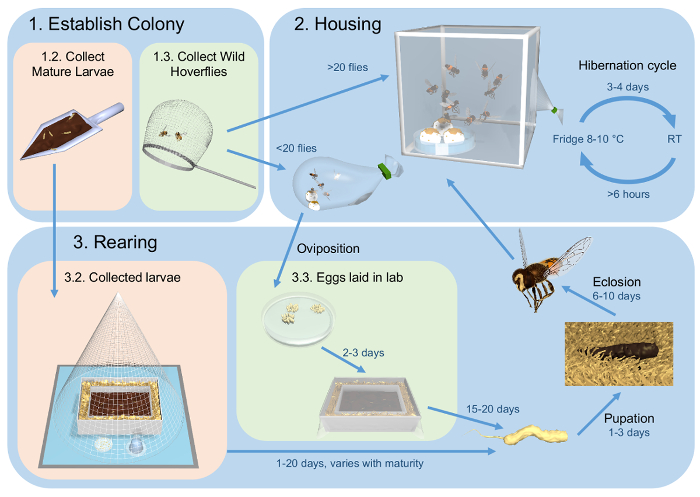
Figure 1: Flow diagram outlining the methods for maintenance of healthy captive population of E. tenax. (1) The procedure described here starts with collection of either mature larvae from cow dung (Step 1.2) or freely flying hoverflies (Step 1.3). (2) The hoverflies are housed in insect cages or plastic bags, dependent on numbers. They are kept in an artificial hibernation cycle at 8-10 °C, which is broken every 3-4 days. (3) Collected larvae are kept in their cow dung (Step 3.2). Eggs laid in the lab are placed in a rabbit feces slurry (Step 3.3). When reaching maturity, 3rd instar larvae crawl into the surrounding saw dust where they pupate. Eclosion occurs after 6-10 days, and the eclosed hoverflies are placed in the housing (Step 2). Please click here to download this file.
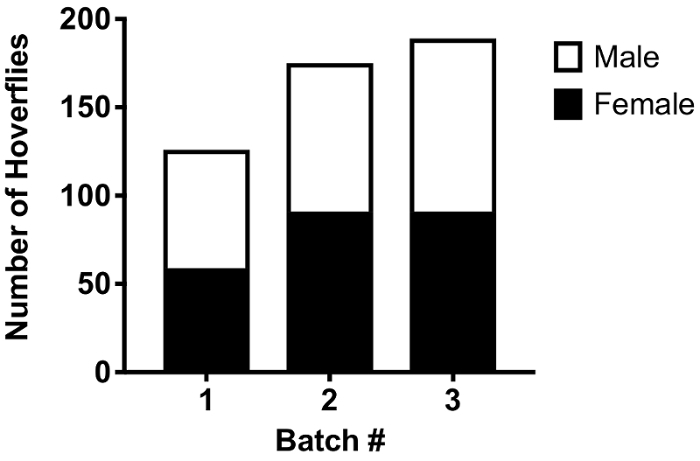
Figure 2: Number and sex ratio of E. tenax successfully reared from individual egg batches. The data show the numbers of E. tenax that eclosed from pupae developed from 3 batches of eggs laid in our lab. The eggs were laid by wild-caught females. The data are color coded for the sex of the flies. There is no significant difference in the ratio.
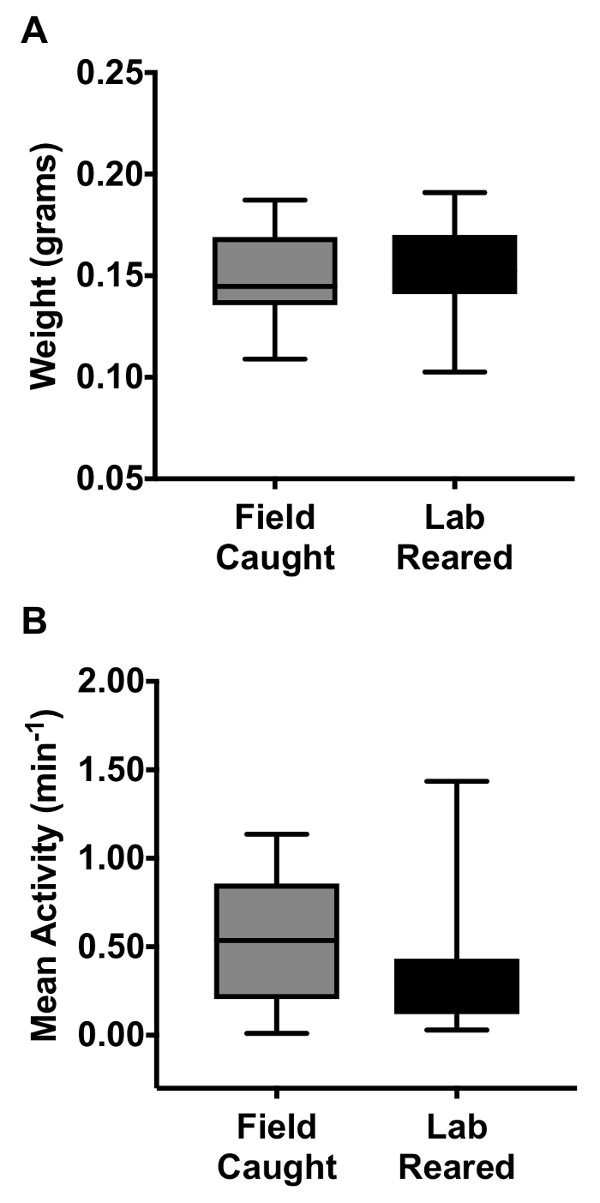
Figure 3: Assessment of population health in laboratory reared and field collected female hoverflies after long term captivity. (A) Weight comparison between lab reared and field captured female hoverflies after 4 months in captivity under artificial hibernation (N=12). (B) Activity levels of lab-reared and field captured female hoverflies after 4 months in captivity under artificial hibernation. The hoverflies' locomotor activity was measured in a Locomotor Activity System by them breaking an infrared beam during movement. As previously, we averaged the activity across 6.7 hours in the middle of the day, on the second full day in the Locomotor Activity Monitor System39 (Nfield caught=9, Nlab reared=12). The central mark of each boxplot shows the median, the edges of the box the 25th to 75th percentiles of the data, and the whiskers extend from the minimum to the maximum of the data.
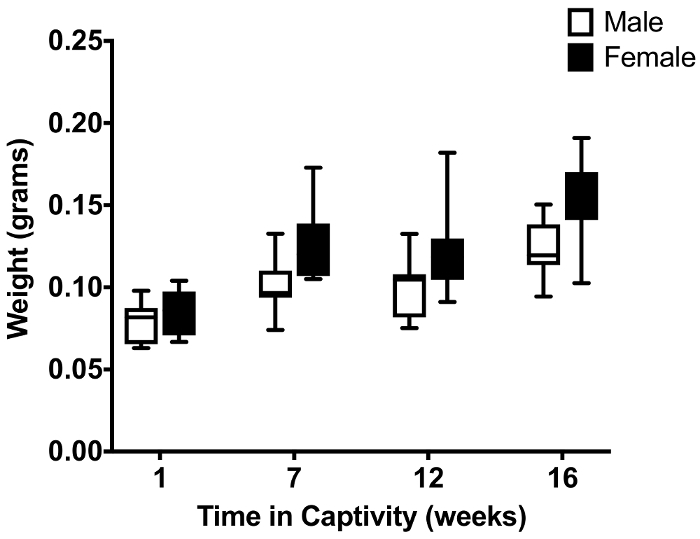
Figure 4: Comparison of the effect of long term maintenance on the weight of lab reared hoverflies of either sex. The data show the hoverfly weight as a function of time kept in captivity under artificial hibernation. As all hoverflies were reared from eggs laid in our lab (N=12 at each data point) and t=0 equals time of pupal hatching, the time in captivity is the same as the age of the animals. The central mark of each boxplot shows the median, the edges of the box the 25th to 75thpercentiles of the data, and the whiskers extend from the minimum to the maximum of the data.

{kind=link}