V. cholerae colonization of zebrafish intestinal tracts
To provide an example of the typical colonization levels we observe, we inoculated 5 x 106 CFU of the pandemic EL Tor V. cholerae strain N16961 in 200 mL of water in a beaker containing several zebrafish. After 6 h of infection, the fish were washed in fresh water and transferred into a beaker of 200 mL of autoclaved infection water as described in the protocol. 18 h after the transfer (or 24 h after the primary infection), the fish were euthanized, and their intestines were taken to determine the colonization levels. Approximately 105 to 106 V. cholerae cells per fish intestine were typically observed in the intestinal tract 24 h post-infection (hpi) (Figure 1) with the 5 x 106 inoculum size.
Quantification of fish diarrhea
Diarrhea was quantified using the four simple assays described above. The first assay, the CFU of excreted V. cholerae, is shown in Figure 1. Serial dilutions of the water 24 hpi were plated to count the V. cholerae CFU. Relatively high levels of excreted V. cholerae are usually observed; in this example, approximately 105 V. cholerae per mL of water were detected. Uninfected fish do not produce detectible V. cholerae (data not shown).
The second assay used to quantify diarrhea is excreted mucin. Figure 2 shows the effects of three different V. cholerae infectious doses on the mucin levels in water 24 hpi. A higher infectious dose is correlated with a higher excretion of mucin into the water. As a control, four fish were placed in an identical beaker of water, but PBS was added instead of the V. cholerae suspension. The control fish excreted very little mucin.
The third diarrheal quantification assay is the OD600 of the water 24 hpi. As shown in Figure 3, the OD600 of this water was significantly higher in the beaker containing zebrafish infected with V. cholerae than in the beaker containing uninfected zebrafish.
Finally, the total protein levels were measured in the water. Water containing the infected fish contained total protein levels nearly twice that observed in the water containing the uninfected fish (Figure 4). Collectively, these four assays illustrate the effects V. cholerae infection has on zebrafish excretion.
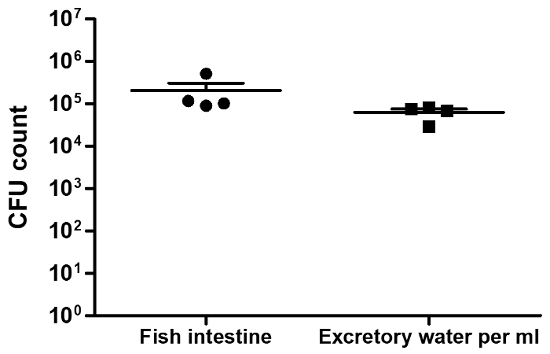
Figure 1: Intestinal colonization of zebrafish by V. cholerae. The fish were infected for 6 h, then washed and incubated for a total of 24 h. The left side of the figure illustrates the total CFU per intestine of four fish (black dots.) The right side of the figure indicates the V. cholerae CFU per mL measured in water 24 hpi (black squares.) The means of both data sets are shown with a large horizontal line and the standard deviation are indicated by error bars. Please click here to view a larger version of this figure.
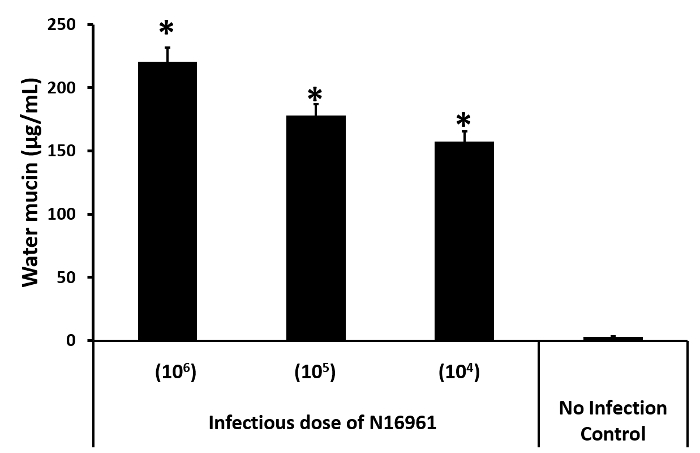
Figure 2: Mucin assay. Three different V. cholerae infectious doses were used, along with an uninfected control, as indicated under the x-axis. The mean values of the mucin detected in the water by the modified Periodic acid Schiff (PAS) assay are indicated by the black bars above each infectious dose. The error bars indicate the standard deviation. The asterisks indicate p <0.05 as determined by Student's unpaired t-test. Please click here to view a larger version of this figure.
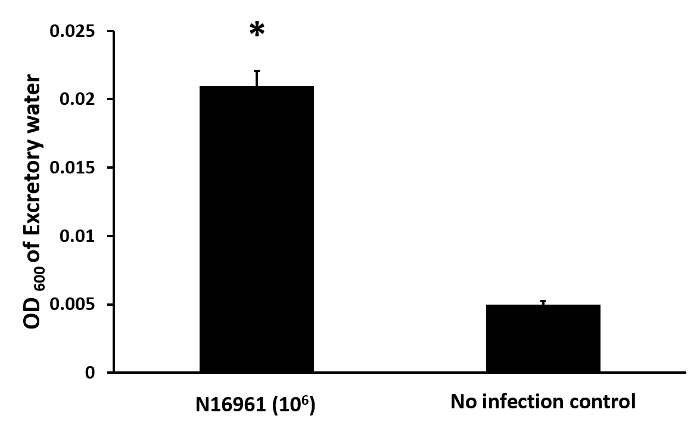
Figure 3: OD600 assay as a proxy for diarrhea. The OD600 of 1 mL of water from the beakers containing either V. cholerae infected or uninfected fish was measured. The black bars indicate the mean value and the error bars indicate the standard deviation. The asterisks indicate p <0.05 as determined by Student's unpaired t-test. Please click here to view a larger version of this figure.
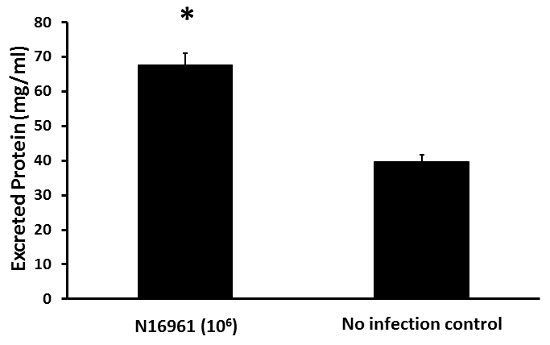
Figure 4: Total protein levels in water. The total protein was estimated by a Bradford assay as described. The black bars indicate the mean values and the error bars indicate the standard deviation. The asterisks indicate p <0.05 as determined by Student's unpaired t-test. Please click here to view a larger version of this figure.
