This section presents examples of results on facial musculature form that can be achieved by using "reverse dissection" in concert with DiceCT scanning. By using "reverse dissection" to create a facial mask, a fuller representation of mimetic (facial) muscle can sometimes be seen than in traditional dissection methodology. This method works on a range of body sizes from the tiny, small-bodied primates, for example the common marmoset Callithrix jacchus (Figure 4), to large-bodied primates such as the chimpanzee Pan troglodytes (Figure 5), and a medium-sized primate such as the rhesus macaque Macaca mulatta (Figure 6). Traditional dissection methodologies may work well on large-bodied primates that have robust mimetic musculature. However, traditional "front approach" dissection methods may not work well with small-bodied primates that have gracile facial muscles. In these cases, some of the facial musculature may be indistinguishable from the surrounding connective tissue and may be lost during dissection.
The iodine stain bound to the mimetic musculature and at least some of the scans are of sufficient quality that we can resolve both individual mimetic muscles (Figure 7) as well as individual muscle fascicles (Figure 8) and, for the first time, obtain whole muscle volumes of these gracile muscles. As shown in Figure 7, some of the very small muscles associated with the external ear are clearly visible in the DiceCT scans. It is not uncommon for these muscles to be missed in some reverse dissection procedures, perhaps due to their small size.
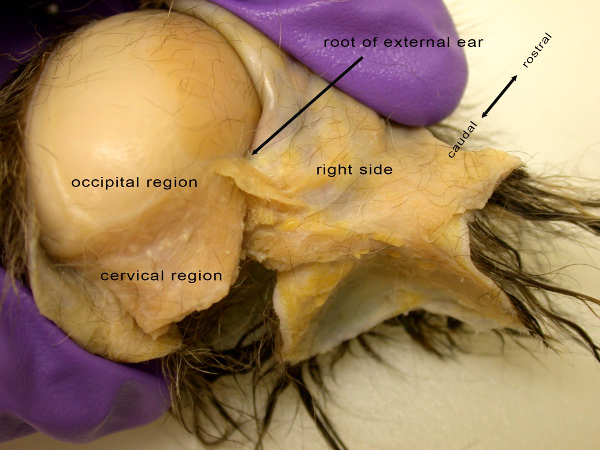
Figure 1: Caudal (or posterior) view of the disarticulated head of a common marmoset (Callithrix jacchus) showing the beginning of the process for creating the facial mask in "reverse dissection". Facial musculature associated with the external ear is shown on the right side of the developing facial mask in shades of orange. Adipose tissue, or fat, is clustered around the musculature in shades of bright yellow. Please click here to view a larger version of this figure.
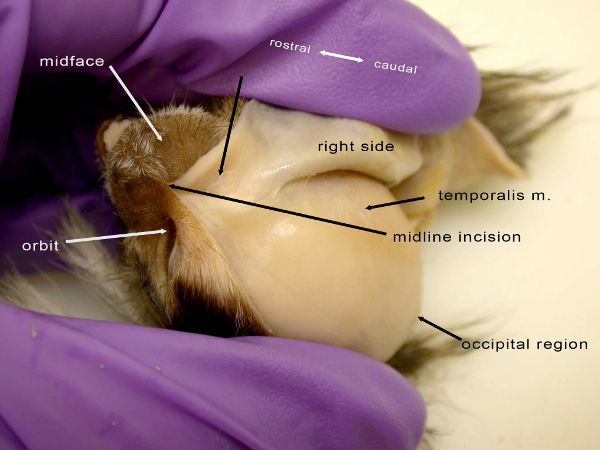
Figure 2: Dorsal view of the disarticulated head of a common marmoset (C. jacchus) showing the middle phase of creating the facial mask in reverse dissection, here removing the mask from the orbital region of the skull. The unlabeled black arrow indicates the area where muscles such as the orbicularis oculi muscle are located, prior to removal of connective tissue. The temporalis muscle is not a facial muscle but is indicated to give an idea of relative location. Please click here to view a larger version of this figure.
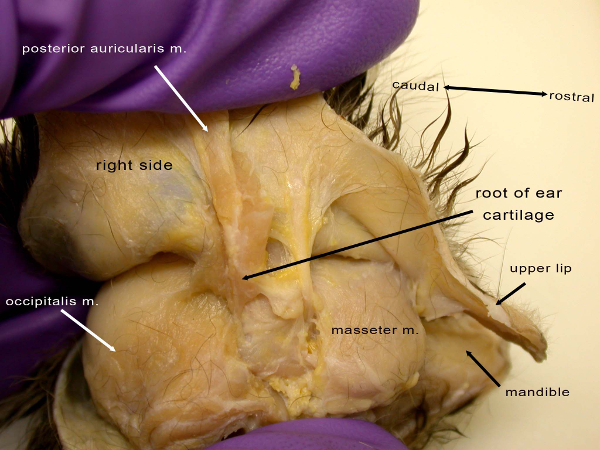
Figure 3: View of the right side of the common marmoset (C. jacchus) showing near the end phase of creating the facial mask in reverse dissection, here removing the mask from the upper and lower lip region of the skull. The masseter muscle is not a facial muscle but is indicated to give an idea of relative location. Please click here to view a larger version of this figure.
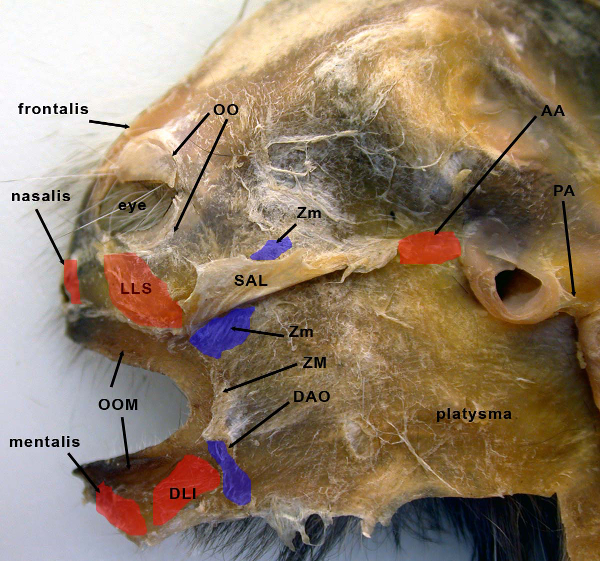
Figure 4: Deep (or inside) view of the entire right side of the facial mask from the common marmoset (C. jacchus), showing the fully dissected facial mask with select muscles indicated. Various muscles are highlighted with color to indicate boundaries. Abbreviations: AA – anterior auricularis muscle; DAO – depressor anguli oris muscle; DLI – depressor labii inferioris muscle; LLS – levator labii superioris muscle; OO – orbicularis oculi muscle; OOM – orbicularis oris muscle; PA – posterior auricularis muscle; SAL – superior auriculolabialis muscle; ZM – zygomaticus major muscle; Zm – zygomaticus minor muscle. This image appeared in Burrows, 20082. Please click here to view a larger version of this figure.
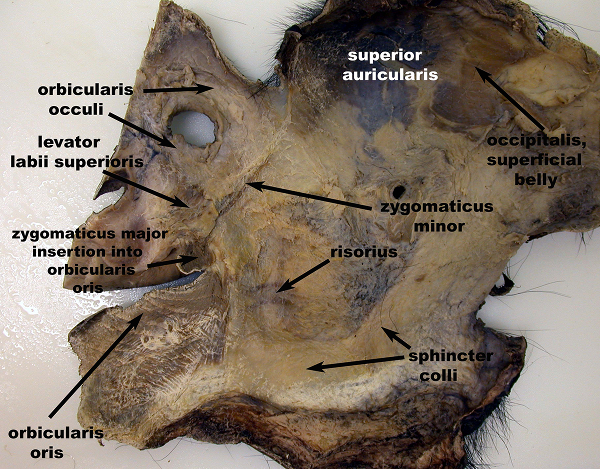
Figure 5: Deep (or inside) view of the entire right side of the facial mask from the common chimpanzee (Pan troglodytes), showing the fully dissected facial mask with select muscles indicated. The risorius muscle is indicated here, a muscle which was previously thought to be present among primates only in humans. This image appeared in Burrows et al., 200615. Please click here to view a larger version of this figure.
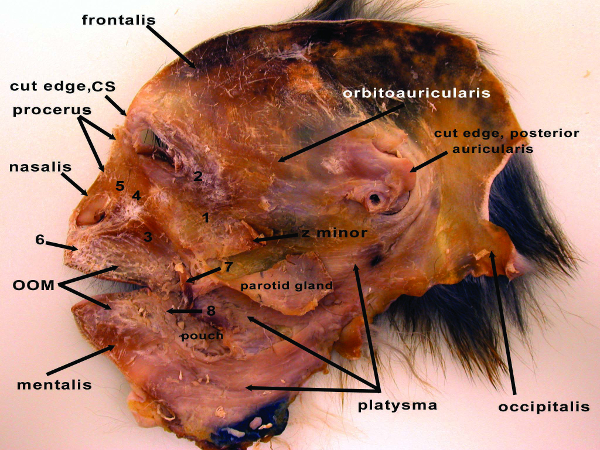
Figure 6: Deep (or inside) view of the entire right side of the facial mask from the rhesus macaque (Macaca mulatta), showing the fully dissected facial mask with select muscles indicated. CS – corrugator supercilli muscle; OOM – orbicularis oris muscle; z minor – zygomaticus minor muscle; 1 – zygomaticus major muscle; 2 – orbicularis oculi muscle; 3 – caninus muscle; 4 – levator labii superioris muscle; 5 – levator labii superioris alaeque nasi muscle; 6 – depressor septi muscle; 7 – cut edge of buccinators muscle; 8 – depressor labii inferioris muscle. This image appeared in Burrows et al., 200916. Please click here to view a larger version of this figure.
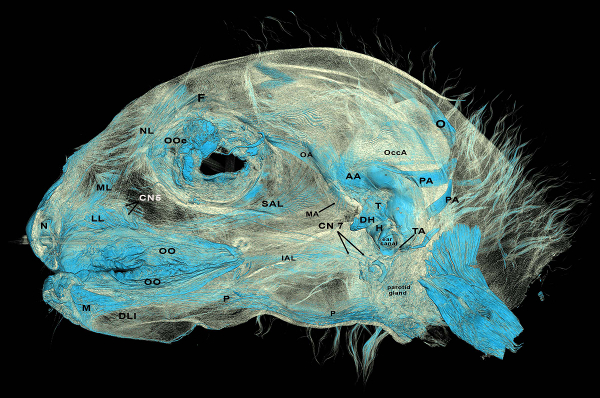
Figure 7: Deep (or inside) view of the entire right side of a DiceCT scan from a Eulemur flavifrons demonstrating the abilities of DiceCT to pick up mimetic muscle fibers. AA: anterior auricularis muscle; CN5: cranial nerve 5; CN7: cranial nerve 7; DH: depressor helicis muscle; DLI: depressor labii inferioris muscle; F: frontalis portion of occipitofrontalis muscle; H: helicis muscle; IAL: inferior auriculolabialis muscle; LL: levator labialis muscle; M: mentalis muscle; MA: mandibuloauricularis muscle; ML: maxillolabialis muscle; N: nasalis muscle; NL: nasolabialis muscle; O: occipitalis portion of occipitofrontalis muscle; OccA: occipitoauricularis muscle; OO: orbicularis oris muscle; OOc: orbicularis occuli muscle; P: platysma muscle; PA: posterior auricularis muscle; SAL: superior auriculolabialis muscle; T: tragicus muscle; TA: tragoantitragus muscle Please click here to view a larger version of this figure.
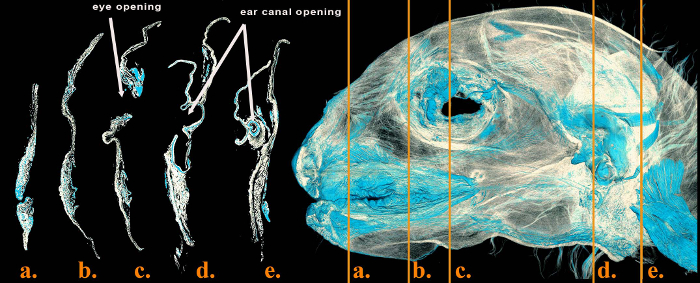
Figure 8: Deep (or inside) view of the entire left side of a DiceCT scan from a Eulemur flavifrons demonstrating serial sections at various points. Deepest blue stain is from areas where there is a heavy presence of mimetic muscle fibers (e.g., around the opening of the external ear, sections a. and b., and the upper orbital region, section c.). Lightest blue stain is from areas where there is less mimetic muscle fiber (e.g., the region of the upper lip, section d.). Please click here to view a larger version of this figure.
