We here described a protocol for the purification of a TAP-tagged chloroplast envelope protein complex from transgenic A. thaliana plants. As shown in Figure 1A, plants expressing the 35S:NTAP-TOC159 construct were used to isolate this complex while plants expressing the 35S:NTAP construct were used as a control. Figure 1B shows detailed steps of the purification of TAP-TOC159 protein complexes followed by mass spectrometry to allow the identification of interacting proteins. The immunoblot analysis (Figure 2, right side) confirms that isolated TAP-TOC159 interacts with TOC75 and TOC33 of the TOC complex. The chloroplast outer membrane kinase KOC1 known to phosphorylate TOC159 also co-isolated with the TAP-TOC159 complex (Figure 2B, right side). The presence of TIC110 reveals that the isolated TAP-TOC159 complex also contains the components of the TIC complex. The FBN1A antibody raised against a plastoglobule protein unrelated to protein import did not recognize the TAP-TOC159 complex indicating the absence of contaminations. In the negative control, immunoprecipitated NTAP protein did not co-isolate with any of the TOC-TIC proteins (Figure 2, left side) and confirms the specificity of the TAP-TOC159 purification.
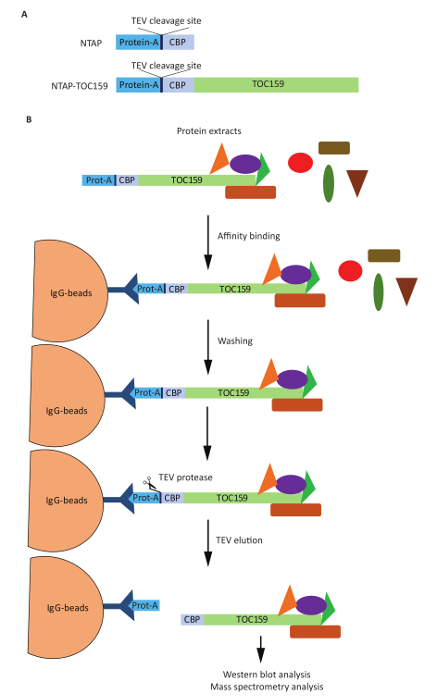
Figure 1. Scheme of the constructs and affinity purification procedure. (A). The 35S:NTAP-TOC159 construct, encoding NTAP-TOC159 was stably expressed in Arabidopsis thaliana. Negative control plants expressed the 35S:NTAP construct, encoding the TAP-tag alone. (B). The step wise affinity purification protocol consists of membrane isolation and solubilization, IgG agarose immuno-purification, washes, TEV protease cleavage and elution. Please click here to view a larger version of this figure.
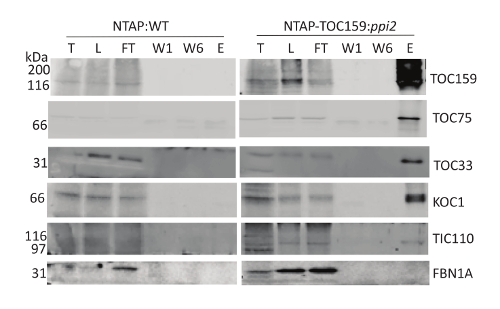
Figure 2. Tandem affinity purification of the TOC159 protein. N-terminally TAP-tagged TOC159 was purified from NTAP-TOC15:ppi2 Arabidopsis plants and TAP protein purified from TAP:WT plants was used as a negative control. Total protein extracts (T), solubilized membrane proteins = load fraction (L), flow through (FT), first and last wash fractions (W1 and W6) and 10% of the TEV eluate were analyzed by Western blotting. The membrane was probed with the antibodies against TOC159 (AT4G02510), TOC75 (AT3G46740), TOC33 (AT1G02280), KOC1 (AT4G32250), TIC110 (AT1G06950), and FBN1A (AT4G04020). Please click here to view a larger version of this figure.
| Table of buffers | ||
| Coupling Buffer | NaHCO3 Adjust to pH 8.5 adjust pH with 0.1 M Na2CO3 |
100 mM |
| Blocking Buffer | Tris–HCl Adjust to pH 8 with HCl. |
100 mM |
| PBS Buffer | Na2HPO4 KH2PO4 NaCl KCl Adjust to pH 7.3 with HCl. |
4.3 mM 1.4 mM 137 2.7 |
| NaCl coupling Buffer | Coupling buffer NaCl |
1 M |
| 2X grinding Buffer | Tris-HCl, pH 7.5 with HCl. NaCl NaF PMSF Plant protease inhibitor cocktail |
100 mM 200 mM 5 mM 1 mM 0.20% |
| 4X solubilisation solution | Glycerol Triton-X100 |
40% 3% |
| Buffer A | 2X grinding Buffer 1X grinding Buffer 4X solubilisation solution |
100 ml 50 ml 25 ml 25% |
| TEV elution Buffer | Tris-HCl, pH 8 with HCl EDTA, pH 8 with HCl NaCl Glycerol Triton-X100 DTT |
50 mM 0.5 mM 100 mM 10% 0.75% 1 mM |
| 2x SDS-PAGE loading Buffer | Tris-HCl pH 6.8 with HCl SDS Glycerol Bromophenol blue DTT |
0.1 M 4% 20% 0.20% 0.2 M |
Table 1. Buffer recipes.
