A schematic diagram of the experimental procedure is provided in Figure 1. The underlying basis of the technique is the unusual ability of CK2 to use GTP for phosphoryl group transfer. Addition of exogenous CK2 holoenzyme along with the GTP analogue, GTPγS, to a cell lysate results in thiophosphorylation of endogenous CK2 substrates. Subsequent treatment of the lysate with the alkylating reagent p-nitrobenzyl mesylate (PNBM) generates a thiophosphate ester moiety on these specific substrate proteins that can then be immunoprecipitated using an anti-thiophosphate ester antibody and ultimately identified by mass spectrometry. Figure 2 depicts a positive result following the addition of CK2 and GTPγS and then PNBM to T98G (glioblastoma) cell lysate. These results demonstrate that CK2-dependent thiophosphorylation and subsequent alkylation were successful. As expected, an enhanced anti-thiophosphate ester signal by Western blotting is observed only in the lane containing the complete kinase reaction and not in the GTPγS only- and PNBM only-treated samples. Shown in Figure 3 is a Coomassie blue-stained gel of the immunoprecipitated and eluted proteins using isotype control IgG or anti-thiophosphate ester antibodies in the presence or absence of excess CK2 and/or GTPγS. These data also demonstrate a positive result as multiple unique bands are evident only in the anti-thiophosphate ester IP lane in which the lysate was incubated with exogenous CK2 and GTPγS. The band indicated with an asterisk was excised from the gel and submitted for protein identification by mass spectrometry. Figure 4 illustrates representative data obtained by mass spectrometric analysis including protein identification, percent coverage, and number of unique peptides identified per protein within the band. Shown are the top ten hits from the submitted band (Figure 3) and information regarding whether or not the protein has been previously identified as a substrate of CK215,16,17. The identity of one of the known immunoprecipitated CK2 substrates, nucleolin15, was confirmed by SDS-PAGE and immunoblotting of the indicated fractions using an anti-nucleolin antibody.
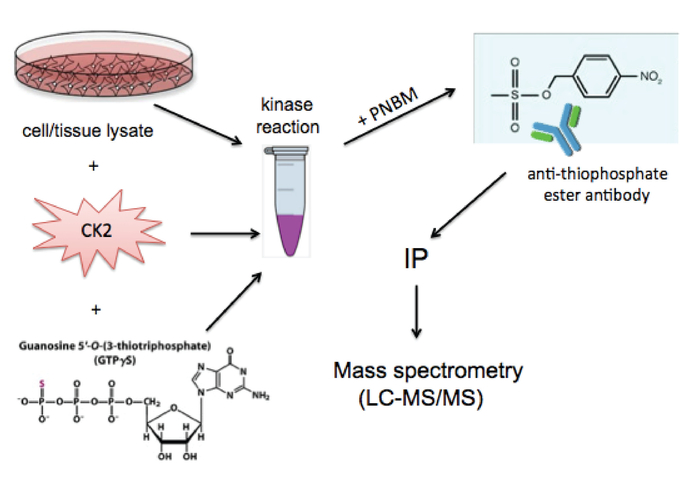
Figure 1: Schematic diagram of the experimental strategy. The GTP analogue, GTPγS, along with excess recombinant CK2 holoenzyme is added to a cell or tissue lysate and allows for thiophosphorylation of substrates by CK2 but not by other endogenous kinases. Thiophosphorylated substrates are next alkylated with PNBM, generating a thiophosphate ester moiety on these proteins, which are then captured via immunoprecipitation (IP) for subsequent identification by liquid chromatography-tandem mass spectrometry (LC-MS/MS). Please click here to view a larger version of this figure.
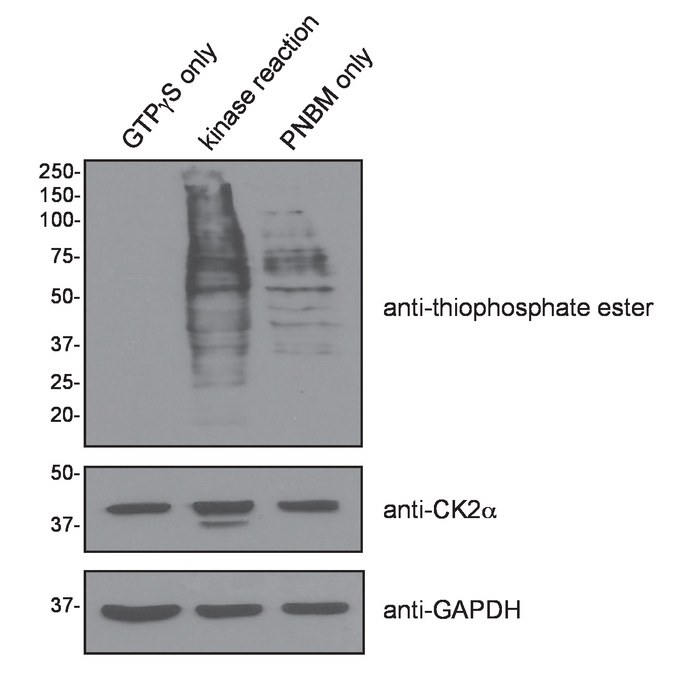
Figure 2: Validation of CK2-dependent thiophosphorylation in whole cell lysate. Whole cell lysates prepared from T98G cells were incubated with GTPγS in the presence (kinase reaction) or absence (GTPγS only) of exogenous recombinant CK2 holoenzyme. PNBM was subsequently added to the indicated reactions, and samples were resolved by SDS-PAGE followed by immunoblotting with the indicated antibodies. Protein molecular weight markers are indicated in kDa. Please click here to view a larger version of this figure.
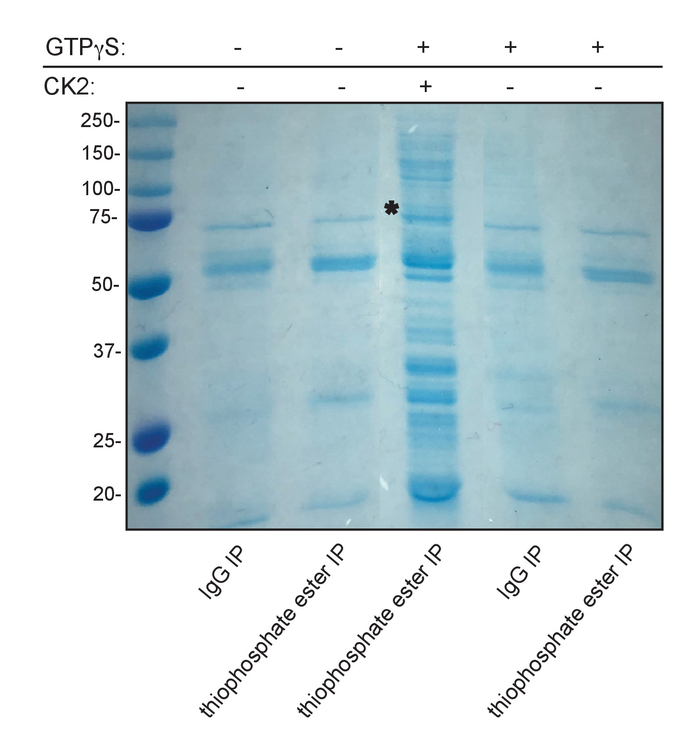
Figure 3: Immunoprecipitation of putative CK2 substrate proteins. Enrichment and visualization of putative CK2 substrates (third lane) is evident as multiple unique bands following immunoprecipitation with anti-thiophosphate ester antibodies. Immunoprecipitates were resolved by SDS-PAGE and the gel was stained with Coomassie blue. The band marked with an asterisk was excised from the gel and submitted for protein identification by mass spectrometry. Protein molecular weight markers are indicated in kDa. IP=immunoprecipitation. Please click here to view a larger version of this figure.
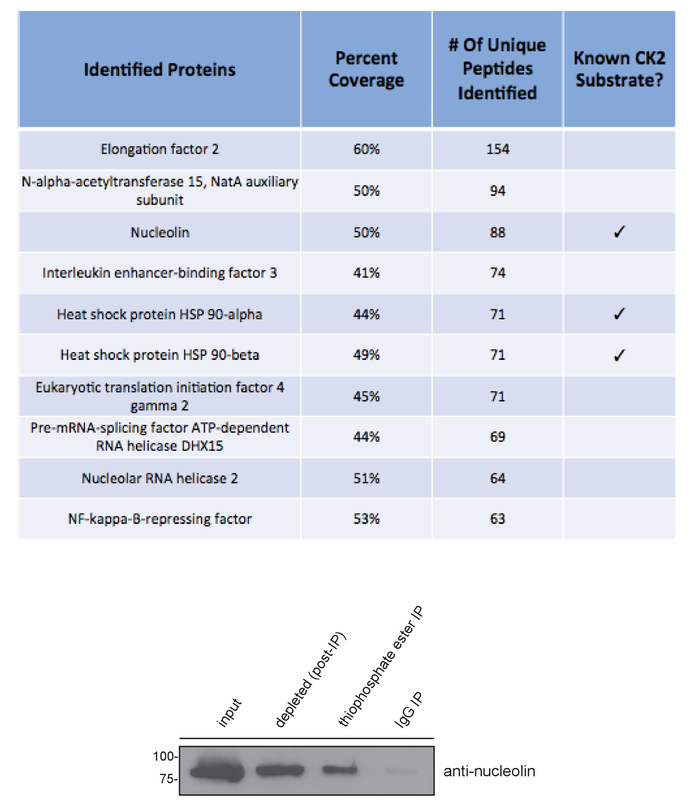
Figure 4: Identification and confirmation of proteins as substrates of CK2 in vitro. Data obtained by mass spectrometry demonstrates that both previously known15,16,17 as well as putative novel CK2 substrates were identified using this experimental approach. Shown are the top ten proteins identified from the excised band (top). The identity of nucleolin, a known CK2 substrate, was confirmed by immunoblotting of the indicated fractions using an anti-nucleolin antibody (bottom). Protein molecular weight markers are indicated in kDa. IP=immunoprecipitation. Please click here to view a larger version of this figure.
| Reagent Stock Concentration | Reagent Final Concentration | Example volumes added based on stock concentrations |
| 1 M Tris pH 7.4 | 20.0 mM | 200 µL |
| 4 M NaCl | 20.0 mM | 50 µL |
| 20% Triton X-100 | 0.50% | 250 µL |
| 1 M MgCl2 | 10.0 mM | 100 µL |
| 1 M DTT | 0.5 mM | 5 µL |
| 200 mM Na3VO4 | 1.0 mM | 50 µL |
| 500 mM NaF | 10.0 mM | 200 µL |
| 500 mM β-glycerol phosphate | 10.0 mM | 200 µL |
| H2O | 8.945 mL | |
| +1 cOmplete Mini tab/10 mL | 1 tablet |
Table 1. Lysis buffer recipe (10 mL, 1X).
